David Linden - Touch
Здесь есть возможность читать онлайн «David Linden - Touch» весь текст электронной книги совершенно бесплатно (целиком полную версию без сокращений). В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Год выпуска: 2014, ISBN: 2014, Издательство: Penguin Books Ltd, Жанр: Психология, Биология, sci_popular, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Touch
- Автор:
- Издательство:Penguin Books Ltd
- Жанр:
- Год:2014
- ISBN:9780241184059
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Touch: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Touch»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Dual-function receptors in our skin make mint feel cool and chili peppers hot. Without the brain’s dedicated centers for emotional touch, an orgasm would feel more like a sneeze—convulsive, but not especially nice. From skin to nerves to brain, the organization of our body’s touch circuits is a complex and often counterintuitive system that affects everything from our social interactions to our general health and development.
In Touch, neuroscientist and bestselling author David J. Linden explores this critical interface between our bodies and the outside world, between ourselves and others. Along the way, he answers such questions as: Why do women have more refined detection with their fingertips than men? Is there a biological basis for the use of acupuncture to relieve pain? How do drugs like Ecstasy heighten and motivate sensual touch? Why can’t we tickle ourselves? Linking biology and behavioral science, Touch offers an entertaining and enlightening answer to how we feel in every sense of the word.
Touch — читать онлайн бесплатно полную книгу (весь текст) целиком
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Touch», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
If the TRPM8 and TRPV1 activation thresholds for detecting cooling and warming are truly designed to help us maintain our core temperatures, then we should expect these thresholds to be different in animals with core temperatures unlike our own. Indeed, when DNA encoding TRPM8 from a chicken, a rat, and a frog (the clawed frog Xenopus laevis ) was used to artificially express TRPM8 channels, it was shown that chicken TRPM8 was tuned to have a warmer threshold of activation: around 86°F, befitting defense of its core temperature of 107°F. The frog, which is not homeothermic and hence needs only to sense extreme cold, has its TRPM8 tuned cooler, activating only at temperatures below 66°F (figure 5.5). 26On the heat-sensing side, thresholds also appear to be set by core temperature. For example, human TRPV1 is activated at temperatures above 109°F, but the zebra fish, which is not homeothermic, has TRPV1 that activates at about 91°F. The bottom line: The thresholds for hot and cold detection in various animals are not random. Rather, they make sense in terms of the temperature regulation each animal needs to achieve in order to function physiologically. 19
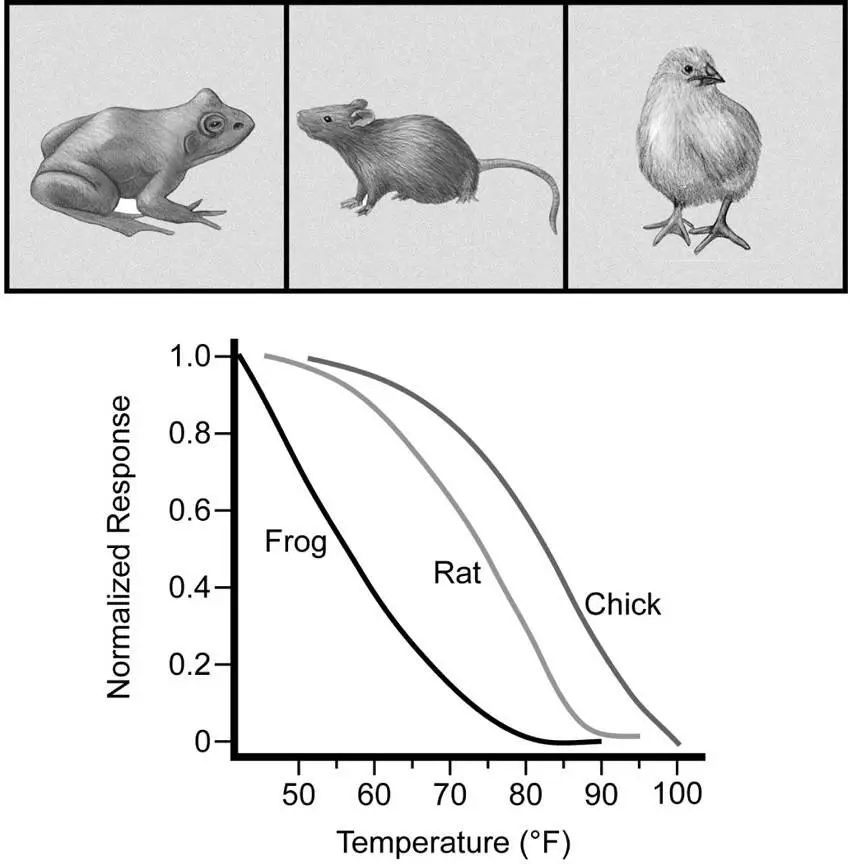
Figure 5.5The threshold for TRPM8 response to cooling is correlated with core body temperature. These temperature response curves were generated through artificial expression of TRPM8 from a frog, a rat, and a chicken. Adapted from B. R. Myers, Y. M. Sigal, and D. Julius, “Evolution of thermal response properties in a cold-activated TRP channel,” PLOS One 4 (2009): e5741, an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.

Years ago, I went on a family visit to my former sister-in-law’s house in Ohio in the late fall. When I awoke in the basement guest room, it seemed more than a little chilly. After bundling up, I headed upstairs and checked the thermostat, which was set to 52°F. That was just right for her, for she was standing happily in the kitchen in shorts and a T-shirt. The following year I made sure to bring along a portable space heater for the guest room. I have another friend who heats his house to the point where the doorframes start to warp, and you almost expect to see the sun-bleached skull of a coyote lying on the hardwood floor.
What accounts for such extremes in individual temperature preference in humans? We know that people with a thinner layer of body fat tend to prefer warmer temperatures, which makes sense in terms of core temperature regulation. We also know that those who are more physically active, even if that activity is mere fidgeting, produce more heat through muscular contraction and thereby prefer cooler temperatures. This may partly explain why young children and teenagers are often resistant to putting on a coat. There is also a daily cyclic variation in core body temperature, and hence external temperature preference. But are there actual differences in the temperature-sensing molecules or circuits of the skin and brain that might explain some of these individual variations? Do temperature preferences run in families? At present, the best answer to these questions is maybe.
We know that different species of animal can have TRPV1 and TRPM8 variants that are tuned to different temperatures. It has also been shown in both rats and humans that drugs that block TRPV1 can produce hyperthermia, a rise in core body temperature, and that drugs that activate TRPM8 can do the same. There is also a rare recessive mutation in humans called WNK1/HSN2. Two copies of this mutant gene produce severe degeneration of sensory neurons, but single-copy carriers of the mutation are unaffected. However, careful measurement of heat and cool detection thresholds showed that the WNK1/HSN2 carriers have their warm threshold shifted slightly cooler and their cold threshold shifted slightly warmer than age- and sex-matched controls. 27But the simple result that one might expect—that genetic variation in human TRPV1 or TRPM8 could account for a portion of individual temperature preference—has yet to emerge. I strongly suspect that my former sister-in-law has a froglike TRPM8 gene, but that has yet to be confirmed.

CHAPTER SIX
Pain and Emotion
On his fourteenth birthday, eager to perform an especially daring trick to impress his friends, a boy jumped off the roof of his family’s house in Lahore, Pakistan. On landing, he got up off the ground and said that he was okay, but died the following day from massive internal bleeding. Despite his major injuries, he never indicated that he was hurt, so his family didn’t take him to the doctor. Not surprisingly, this was no ordinary boy. He was well known as a street performer who would insert knives through his arms and stand on burning coals. People in his neighborhood said that he was fearless because he could feel no pain at all.
Although the boy died before he could be carefully examined, subsequent investigations by Geoffrey Woods, a geneticist at Addenbrooke’s Hospital in Cambridge, England, identified six additional cases in which the ability to sense pain was entirely absent from birth. 1All were children from families of the Qureshi birdari clan in rural northern Pakistan, but because the condition is the result of a rare and random genetic mutation, it can occur anywhere in the world. 2The Pakistani families that Woods studied had emigrated to England and sometimes practiced cousin-marriage. None of the six children had felt any type of pain at any time during their lives—not in their skin, muscles, bones, or viscera. They rarely cried as babies. It’s not that they felt pain but were indifferent to it; rather, they did not experience pain at all. Neurological exams revealed that the children had normal touch perception of fine mechanical stimuli (vibration, pressure, texture), gentle warmth and cooling (but not painful extremes of temperature), tickling, and caressing. If they were to accidentally hit themselves on the thumb with a hammer, they would feel the pressure of the blow, but no pain from it. The injured thumb would then swell, but it wouldn’t throb or ache.
The deficit of the affected children was remarkably specific to pain perception: They had normal reflexes, unimpaired bowel and bladder control, and no obvious disorders of cognition, mood, or social interaction. When asked to describe the meaning of the word pain, none of them could give an appropriate answer, though the older ones had learned what actions would be likely to elicit pain in others (to the point where they could convincingly mimic pain after soccer tackles to draw a penalty for the opposing player). Importantly, the loss of physical pain hadn’t diminished the ability of these children to experience emotional pain. Their feelings could still be hurt, even when their bodies could not. They also appeared to have normal empathy for the emotional pain of others.
Although one might imagine that a life without pain would be idyllic, that turns out not to be the case. Pain occurs in response to stimuli that produce tissue damage. Without it, we do not learn to recoil from sharp blades, boiling liquids, or damaging chemicals. People with congenital total insensitivity to pain are constantly injuring themselves. They bite their own tongues, break their own bones, wear down their joints, and scar their corneas by unknowingly rubbing grit into their eyes. Many do not survive beyond their teenage years. Most people with this condition do not die in a dramatic fashion, like the boy in Pakistan who jumped off the roof. Rather, death is often the result of mundane tissue damage: the ill-fitting shoes that damage the feet, the burning food that scars the esophagus, or even the underwear that is too tight and cuts into the skin of the abdomen. Bacterial infection following such injuries is a constant threat.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Touch»
Представляем Вашему вниманию похожие книги на «Touch» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Touch» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.