David Linden - Touch
Здесь есть возможность читать онлайн «David Linden - Touch» весь текст электронной книги совершенно бесплатно (целиком полную версию без сокращений). В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Год выпуска: 2014, ISBN: 2014, Издательство: Penguin Books Ltd, Жанр: Психология, Биология, sci_popular, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Touch
- Автор:
- Издательство:Penguin Books Ltd
- Жанр:
- Год:2014
- ISBN:9780241184059
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Touch: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Touch»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Dual-function receptors in our skin make mint feel cool and chili peppers hot. Without the brain’s dedicated centers for emotional touch, an orgasm would feel more like a sneeze—convulsive, but not especially nice. From skin to nerves to brain, the organization of our body’s touch circuits is a complex and often counterintuitive system that affects everything from our social interactions to our general health and development.
In Touch, neuroscientist and bestselling author David J. Linden explores this critical interface between our bodies and the outside world, between ourselves and others. Along the way, he answers such questions as: Why do women have more refined detection with their fingertips than men? Is there a biological basis for the use of acupuncture to relieve pain? How do drugs like Ecstasy heighten and motivate sensual touch? Why can’t we tickle ourselves? Linking biology and behavioral science, Touch offers an entertaining and enlightening answer to how we feel in every sense of the word.
Touch — читать онлайн бесплатно полную книгу (весь текст) целиком
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Touch», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Serial complexity appears to be a common theme in the sensory portions of the brain. The visual system, for example, builds recognition of complex visual objects, like faces, from initially primitive features, like spots and bars.
Parallel processing segregates complex touch information into different streams for action. Ultimately, representing the tactile world in the brain is in service of achieving some particular outcome: making a decision, forming a memory, or initiating an action. Heavily processed information flowing out of the higher somatosensory cortex segregates into two different streams. One stream, coursing through a brain region called the insula, informs emotional responses, homeostasis, and some other functions. The insula is now known to be critical for the perceived sense of self. The other stream flows through a region called the posterior parietal cortex and is primarily involved in integrating touch data with information from other senses in order to plan, execute, and fine-tune movements, including the manipulation of objects. 45
While the primary somatosensory cortex mostly responds reliably and stereotypically to touch information, the brain’s higher touch centers are more strongly influenced by cognitive factors like attention, context, motivation, and expectations. We’ll return to these regions in later chapters when we consider the higher cognitive aspects of touch.

I’ve discussed how the distorted nature of the human brain’s touch maps, with larger areas representing body parts like fingers, lips, and feet, reflects the density of touch receptors in particular regions of the skin. But there’s another important factor involved: Touch maps are not permanently defined for one’s entire adult life, but can be changed as a result of individual sensory experience. One nice example of this is found in serious violin, viola, and cello players, professional or semiprofessional, who play for at least twelve hours per week. For these instruments the digits of the left hand are continuously engaged in fingering the strings and producing vibrato effects, a task that involves both enhanced tactile stimulation and exquisite manual dexterity. The right hand, used for bowing, requires much less individual finger movement and tactile feedback. When string players were placed in brain scanners to measure the representation of their hands in the primary somatosensory cortex, it was found that the touch map for the left-hand digits was significantly larger than for the right-hand digits, by about 1.8-fold. (The right- and left-hand touch maps of age-matched nonmusician control subjects, meanwhile, were similar in size.) These same basic results have now been found in three studies performed in different laboratories using somewhat different methods, so the phenomenon appears robust. 46The interpretation of them, however, is less straightforward. The simplest explanation is that years of violin or cello practice have produced an enlargement of the left hand’s territory in the touch map. Another suggestion is that people who happen to be born with or develop unusually large left-hand representations early in life are more likely to take up string instruments and succeed as players. In the same way that children tend to gravitate to sports for which they have a particular natural ability, perhaps they likewise choose an instrument in part based upon some sense of their innate sensory-motor abilities.
To test these two explanations, you’d need to measure the touch maps of the hands before and after musical training. Of course, becoming an accomplished string player takes years, which makes it difficult to perform such a before-and-after type of study. Are there tactile experiences that produce changes in the touch map more quickly? Moving out of the human realm, one striking example is the effect of nursing on first-time mother rats. Norway rats in the lab give birth to litters of eight to twelve pups. In the first few days postpartum, they spend about 80 percent of their time nursing with their twelve nipples, distributed in two rows on their ventral (lower) body surface. When measured twelve to nineteen days after birth, the ventrum portion of the primary touch map of a lactating mother rat was about 1.6-fold larger than that of control rats. (Control rats were either age-matched virgin female rats or nonlactating first-time mothers who had their pups removed at birth.) Fifteen to thirty days after weaning the pups, the ventrum touch map on the lactating mothers had shrunk back to prepregnancy size. These findings suggest that enhanced sensory experience can indeed produce dynamic changes in the touch map, which, at least in some situations, can occur in a matter of days rather than years. 47
As Tom Waits said, “The large print giveth and the small print taketh away.” Experience-dependent plasticity of the touch map works in both directions: Not only can enhanced tactile stimulation cause map expansion, but reduced tactile experience can result in map contraction. When adult rats were fitted with a tiny cast to immobilize one foreleg, the area of the deprived forepaw in the primary touch map was found to shrink by about 50 percent after only seven days; the map representation of the spared forepaw didn’t change at all. The authors of this study removed the cast seven days later and checked the touch map again. At that point the deprived forepaw’s representation in the touch map was still shrunken. It’s likely that, given more time, it would have slowly grown back to its normal size, but the experiment to verify that wasn’t done. 48
All of us are engaged in a slow touch-deprivation experiment throughout our adult lives. From about age twenty to eighty, the density of Merkel disks and Meissner’s corpuscles gradually decreases by about threefold (figure 2.10), and fine spatial acuity falls off to approximately the same degree. Does this mean that decreased fine touch sensation in the elderly can be entirely explained by a loss of shallow mechanoreceptors in the skin? Probably not. One clue is that the reduction in spatial acuity is not precisely uniform over the surface of the body: There’s about a 2.5-fold reduction in fingertip sensitivity but about a fourfold reduction in sensitivity on the soles of the feet and the toes. 49An explanation for this difference is that aging is also associated with a reduction in the speed of spike propagation, from about 150 miles per hour to about 110 miles per hour, in the nerve fibers that convey Merkel and Meissner signals to the brain. This slowing of neural impulses could degrade touch information from distant regions like the toes to a greater degree than it would in closer regions, like the hands or lips. Impaired tactile sensation on the soles of the feet and the toes is an important contributing factor to reduced standing and walking stability in the elderly, often resulting in catastrophic falls.
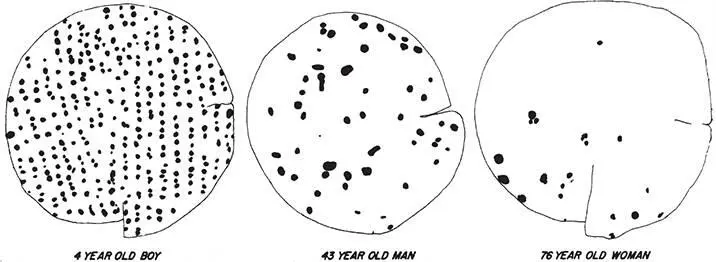
Figure 2.10The density of Meissner’s corpuscles decreases with age in the glabrous skin on the plantar surface of the big toe. These drawings are made from 3-millimeter-diameter punch biopsies. In these samples, the density dropped from 47 to 7 to 3 per square millimeter as age increased. From C. F. Bolton, R. K. Winkelmann, and P. J. Dyck, “A quantitative study of Meissner’s corpuscles in man,” Neurology 16 (1966): 1–9, with permission of Wolters Kluwer Health.
Of course the brain doesn’t just sit there while we grow older. Experience-driven plasticity in the brain may decline a bit with aging, but it never goes away. Our life experience never stops changing our brains. How the primary and higher somatosensory cortices change and adapt in the face of gradually declining mechanosensor density is not presently understood. And we should not assume that if there are plastic changes with aging, they are necessarily beneficial. Cortical plasticity in response to changing touch information might make the problem worse: In subtly rewiring the circuit incorrectly, it might further degrade tactile acuity.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Touch»
Представляем Вашему вниманию похожие книги на «Touch» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Touch» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.