David Linden - Touch
Здесь есть возможность читать онлайн «David Linden - Touch» весь текст электронной книги совершенно бесплатно (целиком полную версию без сокращений). В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Год выпуска: 2014, ISBN: 2014, Издательство: Penguin Books Ltd, Жанр: Психология, Биология, sci_popular, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Touch
- Автор:
- Издательство:Penguin Books Ltd
- Жанр:
- Год:2014
- ISBN:9780241184059
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Touch: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Touch»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Dual-function receptors in our skin make mint feel cool and chili peppers hot. Without the brain’s dedicated centers for emotional touch, an orgasm would feel more like a sneeze—convulsive, but not especially nice. From skin to nerves to brain, the organization of our body’s touch circuits is a complex and often counterintuitive system that affects everything from our social interactions to our general health and development.
In Touch, neuroscientist and bestselling author David J. Linden explores this critical interface between our bodies and the outside world, between ourselves and others. Along the way, he answers such questions as: Why do women have more refined detection with their fingertips than men? Is there a biological basis for the use of acupuncture to relieve pain? How do drugs like Ecstasy heighten and motivate sensual touch? Why can’t we tickle ourselves? Linking biology and behavioral science, Touch offers an entertaining and enlightening answer to how we feel in every sense of the word.
Touch — читать онлайн бесплатно полную книгу (весь текст) целиком
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Touch», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Figure 2.8Touch maps in the primary somatosensory cortex of the human and the star-nosed mole. Left, top: an adult human; middle: the touch map in the human somatosensory cortex; bottom: a drawing of a human with each body part scaled to the size of its representation in the touch map. Note the huge hands, lips, and tongue. Right, top: an adult star-nosed mole; middle: the touch map of the star-nosed mole in the primary somatosensory cortex (shown in a different plane than the human); bottom: a drawing of a star-nosed mole with each body part scaled to its size in the touch map. Note the expansion of the star and forepaws. Illustration by Joan M. K. Tycko. The left top and middle panels are reprinted from David J. Linden, The Accidental Mind: How Brain Evolution Has Given Us Love, Memory, Dreams and God (Cambridge, MA: Harvard/Belknap Press, 2007), 85, with permission of the publisher.
When you dig a little deeper, you discover that there is, in fact, a series of touch maps located in adjacent regions of the cortex. The primary somatosensory cortex in primates can be broken into four different smaller areas, each with its own weirdly distorted map. In addition to receiving information directly from the thalamus, these four areas are highly interconnected with one another. They also send information onward to a set of adjacent areas collectively called the higher somatosensory cortex, which does not receive direct contacts from the thalamus. At present, a total of ten different maps of the body have been found in the somatosensory cortices of the primate brain (four in the primary cortex, six in the higher cortex), and more may yet be revealed.
So where in all of these many interconnected brain maps does the miracle happen? How does our rich, nuanced, and profound sense of touch emerge from this crazily wired system of thinking meat? We consciously perceive that a particular location on the skin has been touched because a particular group of neurons in the cortex is activated. This can occur naturally, through stimulation of the skin, or artificially, through direct brain stimulation, as performed by Penfield. But this is only the first small part of an explanation. The truth is that much remains to be understood about how the brain creates our experience of the tactile world. I will not discuss here all the complex connections within and between these various touch-processing cells and regions of the cortex, but there are a few general principles that are worth knowing.
Information converges from skin to brain. Each neuron in the primary somatosensory cortex ultimately receives converging input from many nerve fibers innervating adjacent regions of the skin. For example, a single nerve fiber running through the arm, conveying Merkel disk information from the fingertip to the spinal cord, will respond to stimulation only in a small spot on the fingertip, about 1 millimeter in diameter. 42But because of this convergent signaling, a Merkel-driven neuron in the finger map of the primary somatosensory cortex is likely to respond to stimulation over a much broader area, about 5 millimeters in diameter. Importantly, this convergent signaling isn’t random and thereby doesn’t merely degrade the resolution of tactile information. Rather, by wiring a particular group of Merkel-driven fibers to the same cortical neuron, one can construct a cortical neuron that responds to a particular touch feature, like a thin bar laid across the finger pad at a particular orientation. Convergent signaling is even more dramatic in areas that are sparsely innervated: A single neuron in the portion of the touch map representing the back can be activated by stimuli ranging over 50 square centimeters of skin, an area about the size of a playing card.
In the primary somatosensory cortex, convergence produces minimal blending of information: Signals from the four mechanosensory receptors are kept largely, but not entirely, separate. Some column-shaped groups of neurons respond preferentially to Merkels, while others respond to Meissners and yet others to Pacinians. So, for example, a single column of cortical tissue, about 0.6 millimeter in diameter, might receive Meissner signals from the pad of the left big toe, while another receives Merkel signals from the right side of the lower lip. 43
Serial processing in the brain extracts increasingly complex touch information. If we examine a block diagram of the areas in the cortex that process touch information, it looks at first glance like a random mess, a bowl of spaghetti (figure 2.9). Upon further examination, however, some themes begin to emerge. There are four areas that comprise the primary somatosensory cortex, all of which receive axons directly from the thalamus, though area 3b receives the lion’s share. By contrast, area 2 gets a bit of information directly from the thalamus but is also activated by all three other primary areas in the neocortex: 3a, 3b, and 1.
If we make recordings of the neurons in area 3b while stimulating the skin, we find that the most effective stimulus to activate them is rather simple, like a thin bar laid across the fingertip at a particular angle. This same simple stimulus will only weakly activate neurons in area 2, however, which respond strongly only to more complex stimuli, such as two- or three-dimensional shapes (like a baseball grasped in the hand). When laboratory animals sustain damage to area 3b, the result is remarkable: The animals become touch-blind, to the point that they seem to be almost entirely unaware of the quality or even the existence of touch stimuli. By contrast, damage to area 2 produces a much more subtle effect: These animals can still detect texture, but their ability to recognize objects by touch alone is lost.
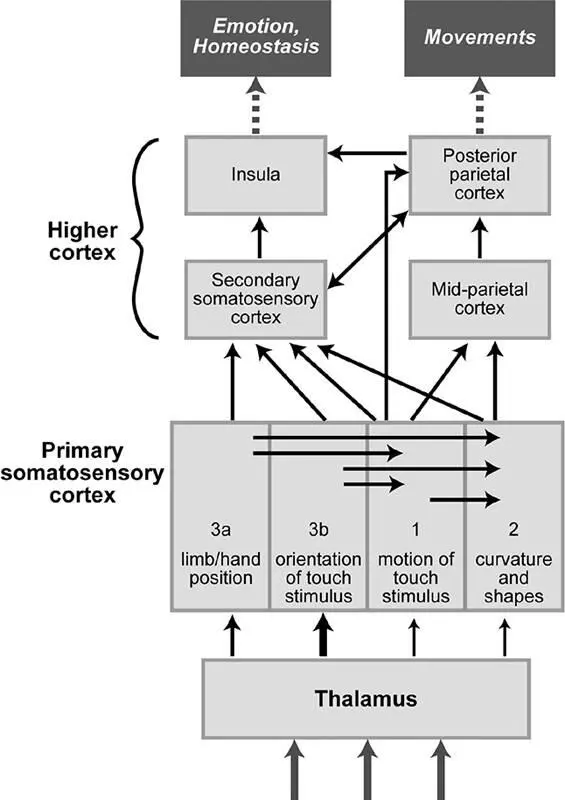
Figure 2.9A simplified wiring diagram of touch-processing regions in the brain. The primary somatosensory cortex receives information directly from the thalamus, performs mostly serial processing to extract different tactile features, and passes the information on to higher somatosensory cortices. The secondary somatosensory cortex is crucial for the recognition of objects and then conveys this information to brain regions involved in emotion and homeostatsis (the insula) and in the early planning of movements (the posterior parietal cortex). Adapted with permission of Professor Steven Hsiao (Johns Hopkins University School of Medicine).
The brain’s wiring diagram for touch-processing regions enables us to make sense of these findings. Area 3b receives basic touch information that is minimally processed. As a result, neurons in this region respond to simple stimuli, as one would predict from a straightforward convergence of axons conveying mechanosensory signals. Because area 3b is a bottleneck for touch information, damage to it is devastating, depriving most of the brain’s downstream processing stations of crucial data. Area 2 has the benefit of receiving not only direct signals from the thalamus but also information from the other primary touch areas, reflecting the processing and computations that those areas perform. As a consequence, area 2 can extract more complex features of touch stimuli, such as object motion, object curvature, and three-dimensional form. When area 2 is damaged, the effects on touch sensation are more subtle, because it is only one of several pathways for information flow to higher areas.
This theme of serial processing with increasing complexity continues as touch information flows farther to the secondary somatosensory cortex. Neurons in this region integrate signals over larger areas (like an entire hand or foot), including regions on both sides of the body. The secondary somatosensory cortex plays an important role in object identification, particularly through exploratory touch. Lesions in this area produce subtle impairments, like loss of the ability to learn a complex object recognition task with one hand and then perform it with the other hand. 44
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Touch»
Представляем Вашему вниманию похожие книги на «Touch» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Touch» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.