Henning Rosenkötter - Motorik und Wahrnehmung im Kindesalter
Здесь есть возможность читать онлайн «Henning Rosenkötter - Motorik und Wahrnehmung im Kindesalter» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на немецком языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Motorik und Wahrnehmung im Kindesalter
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Motorik und Wahrnehmung im Kindesalter: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Motorik und Wahrnehmung im Kindesalter»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Motorik und Wahrnehmung im Kindesalter — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Motorik und Wahrnehmung im Kindesalter», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Nach diesem Blick auf das Gehirn von außen wenden wir uns nun der Feinstruktur des ZNS zu. Beginnen wir mit der Funktion der Nervenzellen, den Neuronen (  Abb. 1.4). Sie erfassen und verarbeiten alle Informationen, die das Gehirn erhält, und sie können gleichzeitig senden und empfangen. Das eingehende Signal kommt entweder über ankommende (afferente) Nervenfasern anderer Neurone oder durch eigene Fasern, die Dendriten. Die Verbindungsstellen (Synapsen) mit anderen Nervenzellen kontaktieren mit ihnen direkt am Zellkörper oder über Synapsen, die auf den Dendriten liegen. Bei manchen Nervenzellen gibt es eine besonders starke und lange auslaufende Faser: das Axon. Eine Erregung im Neuron wandert besonders schnell über das Axon, weil es über Abschnitte verfügt, die Markscheiden genannt werden. Diese Markscheiden-Abschnitte haben Verengungen und Einschnürungen, die Schnürringe.
Abb. 1.4). Sie erfassen und verarbeiten alle Informationen, die das Gehirn erhält, und sie können gleichzeitig senden und empfangen. Das eingehende Signal kommt entweder über ankommende (afferente) Nervenfasern anderer Neurone oder durch eigene Fasern, die Dendriten. Die Verbindungsstellen (Synapsen) mit anderen Nervenzellen kontaktieren mit ihnen direkt am Zellkörper oder über Synapsen, die auf den Dendriten liegen. Bei manchen Nervenzellen gibt es eine besonders starke und lange auslaufende Faser: das Axon. Eine Erregung im Neuron wandert besonders schnell über das Axon, weil es über Abschnitte verfügt, die Markscheiden genannt werden. Diese Markscheiden-Abschnitte haben Verengungen und Einschnürungen, die Schnürringe.
Markscheiden bestehen aus Myelin-Lamellen, die von speziellen Zellen gebildet werden und sich wie Spiralen um die Axone winden. Myelin heißt Mark und ist eine gewundene Membran. Solche markumwickelten Axone können die Erregung schneller leiten als marklose Fasern. Die hohe Übertragungsgeschwindigkeit der markhaltigen (myelinisierten) Fasern kommt dadurch zustande, dass das Myelin wie eine Isolationsschicht wirkt. Dadurch wird die Veränderung der elektrischen Ladung nicht kontinuierlich fortgeleitet, sondern sie springt von einem nicht markumlagerten Schnürring zum nächsten. Myelin bildet sich in der ganzen Kindheit und Jugend und ist der Grund dafür, dass die Erregungsübertragung mit zunehmendem Alter immer schneller wird und das Volumen des Gehirns noch ständig zunimmt, obwohl ab der Geburt keine neuen Neurone mehr gebildet werden.
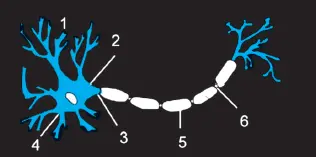
Abb. 1.4: Ein Neuron (1: Dendriten (blau), 2: Zellkörper, 3: Axon, 4: Zellkern, 5: Myelinscheide (weiß), 6: Schnürring)
Die Fortleitung von Signalen im Neuron beruht auf chemischen und elektrischen Vorgängen. Zwischen dem Inneren der Nervenzelle und der Umgebung besteht ein elektrisches Spannungsgefälle, ein elektrisches Potenzial. Diese Spannung kann an der Zellmembran fein abgestuft werden, je nach der Stärke der Erregung des Neurons. Diese wiederum wird von der Stärke der eingehenden Signale bestimmt. Überschreitet das Potenzial an den ausgehenden (efferenten) Fasern eine bestimmte Schwelle, wird plötzlich ein Aktionspotenzial ausgelöst. Die Auslösung folgt dem Alles-oder-Nichts-Prinzip, d. h. entweder ist die Erregung überschwellig und das Potenzial wird ausgelöst oder es wird nicht ausgelöst. Es gibt nur Null (Ruhe) oder Eins (Erregung). Das Aktionspotenzial breitet sich mit großer Geschwindigkeit in den auslaufenden Fasern aus.
Den elektrischen Ruhezustand eines Neurons nennt man Ruhepotenzial. Damit ist gemeint, dass die Zellmembran eine Spannung aufrechterhält, indem ständig durch eine chemische Reaktion Natrium aus der Zelle herausgepumpt und Kalium hineingelassen wird. Diese Natrium-Kalium-Pumpe führt an der Zellmembran zu einem Spannungsungleichgewicht, eben dem Ruhepotenzial. In der Membranwand gibt es Kanäle für Ionen, durch die bei einer bestimmten Spannung zwischen dem Inneren der Zelle und dem Zellaußenraum schlagartig innerhalb einer Millisekunde Natriumionen in das Zellinnere einströmen. Das Ruhepotenzial, eine im Zellinneren negative Ladung, kehrt sich nun plötzlich in eine positive Ladung um. Diesen Potenzialumschwung nennt man ein Aktionspotenzial. Es kann über das Axon an andere Zellen fortgeleitet werden. Das Aktionspotenzial besteht aus einem Entladungsanteil (Depolarisation) und einer kurzen Phase, in der der Natriumeinstrom nach einer Millisekunde abgestoppt und Kalium ausgeschleust wird (Repolarisation), um den ursprünglichen Ruhezustand wiederherzustellen. Nach Ablauf des Aktionspotenzials ist das Neuron für 1–2 Millisekunden nicht wieder erregbar (Refraktärzeit). Von außen kommende Reize können zwar aufgenommen werden, aber sie führen nicht zu einem neuen Aktionspotenzial.
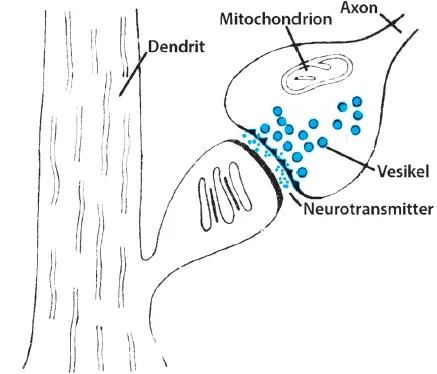
Abb. 1.5: Struktur einer Synapse zwischen Axon und Dendrit
Wenn die Erregung mit einem Aktionspotenzial über ein Axon läuft, wird sie über viele knospenartige Ausläufer (Synapsen) an benachbarte Zellkörper und deren Dendriten weitergegeben. In diesen Synapsenknöpfchen gibt es Bläschen, die prall mit Botenstoffen (Neurotransmittern) gefüllt sind. Ihnen gegenüber liegt an der Synapse des benachbarten Zellkörpers oder Dendriten eine Empfangsmembran. Kommt nun das Aktionspotenzial an die Synapse, werden die Neurotransmitter innerhalb von einer Millisekunde aus den Bläschen entlassen und durch die Synapsenmembran in den Zwischenraum zwischen Synapse und Empfangsmembran (postsynaptische Membran) geschickt ( Abb. 1.5). Den Synapsenspalt überschreiten sie und gelangen an der Empfangsmembran auf spezielle Rezeptoren, die spezifisch immer nur einen bestimmten Botenstoff binden. Die Bindung der Botenstoffe an die Rezeptoren löst in der Nachbarzelle wieder einen Spannungsunterschied aus. Je nach Art des Neurotransmitters und je nach Art des Rezeptors wirkt dieses Potenzial an der Nachbarzelle erregend oder hemmend.
Das einzelne Neuron kann also nach dem Alles-oder-Nichts-Prinzip erregt werden und eine Erregung aussenden oder nicht. Wie kann es dann eine Abstufung der Erregung geben? Es gibt zwei Antworten: Zum einen wirkt sich die Stärke des Reizes auf die Anzahl der Aktionspotenziale pro Zeiteinheit aus. Die andere Antwort ergibt sich, wenn man eine Gruppe von Neuronen betrachtet: Manchmal überwiegen die hemmenden, manchmal die erregenden Impulse. Die Modulation einer Information geschieht durch die Summe von erregenden und hemmenden Einflüssen. Die Feinjustierung ist also in der Zusammenarbeit von funktionell kooperierenden Neuronen möglich.
Eine Sonderform einer synaptischen Verbindung ist die Verbindung zwischen einem Axon und einer Muskelzelle: Die Stelle, an der eine Synapse auf eine Muskelfaser trifft, nennt man die motorische Endplatte. Das ankommende Axon bildet einen synaptischen Endkolben, der viele kleine Energie liefernde Zellorganellen (Mitochondrien) und Bläschen (Vesikel) enthält ( Abb. 1.6).
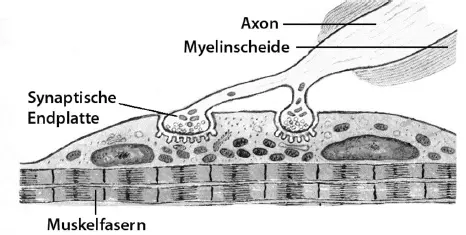
Abb. 1.6: Motorische Endplatte: eine Synapse an der Muskelfaser
Die Vesikel sind vollgestopft mit dem Neurotransmitter Acetylcholin. Kommen Aktionspotenziale über das zuleitende Axon an, so öffnen sich Kalziumionenkanäle. Die acyetylcholinhaltigen Bläschen entleeren sich an der Zellmembran in den synaptischen Spalt. Das freigesetzte Acetylcholin bindet sich an die Rezeptoren der Muskelfaser-Endplatte. Dies ist das Signal zur Kontraktion der darunter liegenden Muskelfaser. Die Größe einer motorischen Einheit entscheidet, wie viele Skelettmuskelfasern sich gleichzeitig zusammenziehen.
Zusammenfassung in Form eines Glossars
1. Makroskopie
Aufbau des Zentralnervensystems
Интервал:
Закладка:
Похожие книги на «Motorik und Wahrnehmung im Kindesalter»
Представляем Вашему вниманию похожие книги на «Motorik und Wahrnehmung im Kindesalter» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Motorik und Wahrnehmung im Kindesalter» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.