Dr. med. Ludwig Manfred Jacob - Prostatakrebs-Kompass
Здесь есть возможность читать онлайн «Dr. med. Ludwig Manfred Jacob - Prostatakrebs-Kompass» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на немецком языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Prostatakrebs-Kompass
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Prostatakrebs-Kompass: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Prostatakrebs-Kompass»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
„Prostatakrebs-Kompass“ mit nahezu 1000 zitierten Studien erklärt die vielen Widersprüche in Ernährungsfragen und liefert ein mehrdimensionales Gesamtbild. Es vereint die epidemiologisch und klinisch besten Konzepte zur Ernährungs- und Lebensweise bei Prostatakrebs in einem praxisnahen Aktionsplan.
Das Buch richtet sich gleichermaßen an Fachleute, die medizinisch oder ernährungswissenschaftlich tätig sind, wie an alle, die sich fundiert über die Zusammenhänge und Ursachen von Prostataerkrankungen informieren und Verantwortung für ihre Gesundheit übernehmen möchten. Die Empfehlungen sind besonders wertvoll für Männer mit familiär erhöhtem Prostatakrebsrisiko, für Männer mit Niedrigrisiko-Prostatakrebs, die sich für eine aktive Überwachung entschieden haben, sowie für Männer, die nach erfolgter Primärtherapie effektiv das Risiko für ein Rezidiv minimieren wollen.
Aus dem Themenspektrum:
• Erst wächst das Bauchfett, dann die Prostata, schließlich oft der Prostatakrebs – Zufall oder Kausalität?
• Welche Rolle spielen Insulin und IGF-1 bei Prostatakrebs, wie senkt man sie?
• Gutartige Prostatavergrößerung und Prostatitis – harmlos oder Risikofaktor?
• Welche Ernährungs- und Lebensweise schafft den Nährboden, welche hemmt das Wachstum von Krebs?
• Welche Lebensmittel sollte man bei Prostatakrebs essen, welche meiden?
• Was ist das Gesundheitsgeheimnis der mediterranen und asiatischen Küche?
• Welche Nahrungsergänzungsmittel nützen, welche schaden?
• Welche Rolle spielen Kanzerogene, Androgene und Östrogene?
• Wie beugt man effektiv Knochenmetastasen vor?
• Psychoonkologie: Je schlechter die Prognose, desto wichtiger die Seele
• DNA-Zytometrie als wertvolle Entscheidungshilfe (Gleason-Grading – Konsens statt Evidenz, Beitrag von Prof. A. Böcking und Josef Dietz)
Mit Geleitwort von Prof. Dr. med. Klippel:
Mit „Prostatakrebs-Kompass“ hat Dr. med. L. M. Jacob ein hochaktuelles, wissenschaftlich fundiertes Buch verfasst, das wichtige, bisher eher weniger bekannte „Wahrheiten“ über das Prostatakarzinom
einer breiten Öffentlichkeit mitteilt und das nicht nur an Ärzte gerichtet ist, sondern auch und gerade an den Prostatakrebs-Patienten. Das vorliegende Buch von Dr. Jacob setzt nicht nur einen neuen Maßstab in der verständlichen Vermittlung aktueller wissenschaftlicher Erkenntnisse, sondern auch in der lebens-praktischen Umsetzung wertvoller Ratschläge für Betroffene. Ein neues Standardwerk“, dessen hoher Anspruch voll und ganz erfüllt wird.
Prostatakrebs-Kompass — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Prostatakrebs-Kompass», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Auch wenn bei Prostatitiden in nur 5 - 10 % der Fälle ein Krankheitserreger ausfindig gemacht werden kann (Weidner et al. , 1991), können entzündliche Infektionskrankheiten in der Vorgeschichte nachweislich eine Rolle spielen: Eine Metaanalyse zeigte, dass Männer, welche zuvor mit Treponema pallidum (2,3-faches Risiko) oder Gonokokken (1,3-faches Risiko) infiziert waren, ein gesteigertes Prostatakarzinomrisiko hatten (Dennis et al. , 2002). Ein vergleichbarer Befund ergab sich für Trichomonaden bei einer Untersuchung an 691 Männern (Sutcliffe et al. , 2006).
Eine PCR-Analyse zur Detektion von bakterieller 16S rRNA ergab eine sehr hohe Korrelation von Prostatitis und der Anwesenheit von Bakterien, was nahelegt, dass diese viel häufiger als diagnostiziert Prostatitis auslösen (Hochreiter et al. , 2000). Wie PCR-Untersuchungen von Prostatakarzinomen auf virale und bakterielle DNA von 83 Erregern ergaben, verteilen sich die Mikroorganismen nicht homogen auf das Prostatagewebe sondern bilden abgegrenzte Herde, welche möglicherweise die Entstehung des späteren Karzinoms, das ja ursprünglich immer aus einem lokalen Zellklon entsteht, begünstigen (Sfanos et al. , 2008). Dies dürfte auch deshalb interessant sein, weil in einer Prostata häufig multiple Karzinome feststellbar sind, die sich auch getrennt voneinander entwickelt haben können.
Erreger kommen häufig über den Harnweg in die Prostata, jedoch lässt die anatomische Nähe zum Mastdarm (s. Abb. 7) auch an eine Einwanderung von Darmbakterien denken. In einer Studie an Mäusen wurde z. B. für die Infektion mit Escherichia coli , einem typischen Darmkeim und Erreger von Harnwegsinfekten, nachgewiesen, dass die Infektion zu atypischen Hyper- und Dysplasien, oxidativen DNA-Schäden und gesteigertem Zellwachstum führte (Elkahwaji et al. , 2007).
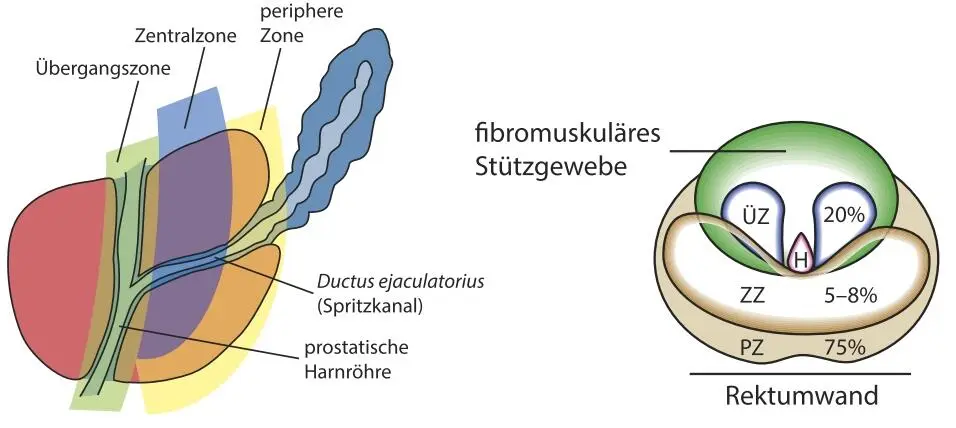
Abb. 7: Lokalisierung der Prostatazonen und Nähe zum Rektum (ÜZ: Übergangszone; H: Harnröhre; ZZ: Zentralzone; PZ: periphere Zone)
3.7.2 Proliferative inflammatorische Atrophie als Präneoplasie
In der peripheren Zone führt die chronische Prostatitis zur postinflammatorischen Atrophie des Drüsenparenchyms. Immunhistochemische Untersuchungen der Gruppe um De Marzo beschreiben die proliferative inflammatorische Atrophie (PIA) (De Marzo et al. , 1999). Dabei handelt es sich um eine entzündlich überlagerte Atrophie der peripheren Zone, die charakteristische Marker aufweist: Überexpression der Gene für Glutathion-S-Transferase (GST) und Cyclooxygenase-2 (COX-2), Überexpression des antiapoptotischen Proteins BCL-2, gesteigerte Proliferationsaktivität und Verlust des Zellzyklusinhibitors p27.
Die Glutathion-S-Transferase ist ein Entgiftungsenzym, das durch entzündliche Prozesse induziert und aktiviert wird. Es gehört übrigens auch zu den Schutzenzymen, dessen Expression ER-beta-vermittelt gesteigert wird (vgl. Kapitel 3.5.2, ab Seite 36) (Montano et al. , 2000 und 2004; Montano und Katzenellenbogen, 1997). In fast allen HGPIN und Prostatakarzinomen geht dieses Enzym durch Hypermethylierung der Promotorregion des GSTP1-Gens verloren. Die Inaktivierung von GSTP1 scheint also eine wichtige Rolle bei der Tumorentstehung in der Prostata zu spielen und der Nachweis im Urin könnte sich demnächst als prognostischer Biomarker eignen (Woodson et al. , 2008). Im Stadium der PIA zeigt die Überexpression von GSTP1 und dem COX-2-Gen, dass diese atrophischen Veränderungen einem chronischen oxidativen Stress ausgesetzt sind. Dabei schützt GSTP1 zunächst das Epithel vor oxidativen Genschäden. Erst bei Hypermethylierung und Inaktivierung der protektiven GSTP1 entsteht dann aus der PIA entweder direkt oder indirekt über HGPIN das Prostatakarzinom (Nelson et al. , 2004).
Im Gegensatz zur protektiven Funktion der GSTP1 ist das COX-2-Gen direkt an der malignen Transformation des Prostataepithels beteiligt und bietet einen möglichen Angriffspunkt für die Chemoprävention. Das COX-2-Gen wird nicht nur in der PIA, sondern auch in einem Teil der HGPIN und der Prostatakarzinome überexprimiert. COX-2-Inhibitoren führen zur Rückbildung von HGPIN im Tiermodell. Einige epidemiologische Studien finden einen Zusammenhang zwischen der chronischen Einnahme von Acetylsalicylsäure und einer geringen Detektionsrate des Prostatakarzinoms (Nelson et al. , 2004). Auch Omega-3-Fettsäuren (Fischöl) vermindern die Expression von COX-2, modulieren den Eicosanoid-Stoffwechsel und bewirken eine Aktivierungshemmung von NF-kappaB, die letztendlich dazu führt, dass neben COX-2 auch andere entzündliche Proteine (z. B. TNF-alpha) vermindert exprimiert werden (Hedelin et al. , 2007; Novak et al. , 2003). Allerdings treten diese Effekte vor allem bei einer sehr fettarmen Ernährung ein. Die Wirkung von Fisch und Fischöl ist komplex und ambivalent und wird daher noch ausführlicher erörtert (s. Kapitel 4.3.8, ab Seite 96).
Polyphenole, insbesondere Granatapfel-Polyphenole, haben eine herausragende antiinflammatorische und antioxidative Wirkung, die eingehend im Kapitel 5.4 (ab Seite 128) behandelt wird.
3.7.3 Proentzündliche Faktoren
Oxidativer Stress durch reaktive Sauerstoff- (ROS) und Stickstoffspezies (RNS) ist ein entscheidendes Bindeglied zwischen Entzündung und Kanzerogenese. Zu den wichtigsten endogenen freien Radikalen gehören ROS wie Superoxid-, Hydroxyl-, Hydroperoxyl-, Peroxyl- und Alkoxy-Radikale sowie RNS wie das Nitroxid- und das Pernitroxid-Radikal. Die Bildung freier Radikale ist zwar einerseits lebenserhaltend im Rahmen der Atmungskette oder als toxische Waffe der Immunabwehr. Andererseits werden freie Radikale jedoch im Rahmen von Entzündungsprozessen verstärkt gebildet und können vor allem im Falle einer Chronifizierung mit Erschöpfung der antioxidativen Schutzsysteme Biomoleküle wie Proteine, Polysaccharide und die DNA angreifen und schädigen.
So fördern freie Radikale zum einen die Tumorinitiation durch ihre direkte DNA-schädigende Wirkung, zum anderen beeinflussen sie unabhängig von ihrer Genotoxizität die weiteren Stadien der Kanzerogenese wie Tumorpromotion und -progression (vgl. Kapitel 3, ab Seite 17), indem sie mit MAPK-Kinase-Signalwegen (Mitogen-aktivierte Proteinkaskade) interagieren (vgl. Abb. 8; Lee und Lee, 2006a). Dies hat folgende Konsequenzen: Die Aktivierung der MAPK-Signalwege aktiviert den proentzündlichen Transkriptionsfaktors NF-kappaB und das Aktivator-Protein-1 (AP-1). AP-1 fördert einerseits direkt die Zellproliferation und Metastasierung und zum anderen induziert es ähnlich wie NF-kappaB die Expression proentzündlicher und prokanzerogener Gene (COX-2, VEGF, Cyclin D1 und MMP-9).
Die Aktivierung von NF-kappaB, die auch direkt durch ROS bewirkt werden kann, erfolgt über eine Phosphorylierung von IkappaB, das im Ruhezustand NF-kappaB daran hindert, in den Zellkern zu dringen und an die DNA zu binden. Nach Phosphorylierung über MAPK-Signalwege löst sich IkappaB aus dem Komplex mit NF-kappaB, das nun seine Funktion als Transkriptionsfaktor erfüllt: NF-kappaB steigert die Transkription proentzündlicher Gene wie Cyclooxygenasen, Interleukine und Matrixmetalloproteasen (MMPs).
Ein weiterer Effekt der veränderten Genexpression durch NF-kappaB ist die Dysregulation der Connexine, welche die Kanäle der GJIC ( Gap-Junction Intercellular Communication ) aufbauen (Lee et al. , 2006). Gap Junctions sind Anhäufungen von Transmembrankanälen, die die Cytoplasmen benachbarter Zellen direkt verbinden. Eine intakte GJIC ist essentiell für die Nährstoffversorgung der Zelle, die Regulation von Wachstum und Differenzierung sowie die Kommunikation mit anderen Zellen und ist bei Krebszellen meist nicht mehr vorhanden. Die Hemmung der GJIC fördert die Tumorpromotion (Lee und Lee, 2006b).
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Prostatakrebs-Kompass»
Представляем Вашему вниманию похожие книги на «Prostatakrebs-Kompass» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Prostatakrebs-Kompass» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.