Tran Quoc Khanh - Beleuchtung in Innenräumen - Human Centric Integrative Lighting
Здесь есть возможность читать онлайн «Tran Quoc Khanh - Beleuchtung in Innenräumen - Human Centric Integrative Lighting» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на немецком языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Beleuchtung in Innenräumen - Human Centric Integrative Lighting
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Beleuchtung in Innenräumen - Human Centric Integrative Lighting: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Beleuchtung in Innenräumen - Human Centric Integrative Lighting»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Beleuchtung in Innenräumen - Human Centric Integrative Lighting — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Beleuchtung in Innenräumen - Human Centric Integrative Lighting», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Lichtmikroskopisch kann man im sog. Corpus geniculatum laterale (CGL) von Primaten sechs Schichten unterscheiden. Die Schichten 2, 3 und 5 erhalten Fasern aus dem ipsilateral gelegenen Auge, die Schichten 1, 4 und 6 aus dem kontralateralen Auge. Innerhalb dieser Schichten bekommen benachbarte Neuronen Signale von jeweils benachbarten Ganglienzellen der Retina. Die Schichten 5 und 6 gehören zu den magnozellulären Schichten. Sie bestehen aus großen Zellen und erhalten ihre Eingangssignale von den sog. Parasolzellen der Retina. Letztere Zellen haben größere Dendritenbäume und somit eine geringere Ortsauflösung, aber eine größere zeitliche Auflösung. Die parvozellulären Schichten 1–4 bestehen aus kleineren Neuronen und bekommen ihre Eingangssignale von sog. Midget-Zellen der Retina. Diese haben kleinere Dendritenbäume, eine hohe Ortsauflösung, aber eine geringere zeitliche Auflösung.
Vom CGL werden die Signale über die breit aufgefächerte Sehstrahlung, teilweise über die sog. Capsula interna , ein weiterer Knotenpunkt großer Nervenbahnen, zur primären Sehrinde im Hinterhauptslappen weitergeleitet.
Der visuelle Kortex : Mit dem visuellen Kortex ist hier der sog. primäre visuelle Kortex gemeint. Er ist unterteilt in zwei Pfade, den sog. ventralen und den sog. dorsalen Pfad. Dem ventralen Pfad werden in der Hirnforschung Form- und Objekterkennung sowie die Speicherung im Langzeitgedächtnis zugeschrieben. Dorsal werden Bewegungen und Orte erkannt sowie Bewegungen des eigenen Körpers koordiniert. Der visuelle Kortex ist in die visuellen Rinden V1–V5 [8] aufgeteilt, denen verschiedene Funktionen zugewiesen werden.
3.3.1.4 Gesamte Netzwerkstruktur des Sehsystems – ein Überblick nach [6]
Fällt die optische Strahlung mit einer bestimmten Intensität und Spektrum in den menschlichen Augenapparat ein, kann man den visuellen Verarbeitungsweg aus der heutigen Sicht der Forschung schematisch, wie in der Abb. 3.9dargestellt, beschreiben.
Nach dem Passieren der Augenoptik (s. Abb. 3.9) trifft die optische Strahlung auf die drei Zapfentypen (L, M, S), Stäbchen R und auf die fotoempfindlichen Ganglienzellen G. Aus den Signalen der Zapfen L, M, S werden die Signale für die beiden Opponenzkanäle (L−M) und S−(L+M) sowie für den Luminanzkanal (L+M) gebildet (s. auch Kap. 2). Diese Bildung findet auf dem Weg zu den „normalen“ Ganglienzellen sowie auch im Kortex statt. Im mesopischen Bereich (wie am Abend bei der Straßenbeleuchtung oder im Kino) kommen die Signale der Stäbchen hinzu [7–10].
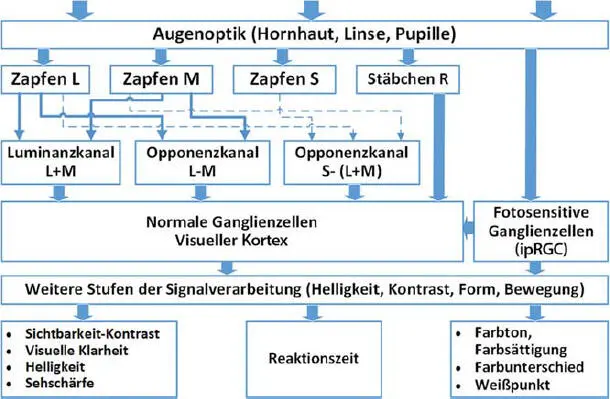
Abb. 3.9 Verarbeitungsschaltung für die visuelle Wahrnehmung. Quelle: TU Darmstadt.
Jüngste Untersuchungen, wie in [11] beschrieben, wiesen die Beteiligung von melanopsinhaltigen Ganglienzellen an der Bildung der Helligkeitswahrnehmung nach, was auch durch die Modellierung der Helligkeit in [12] auf der Basis der psychophysischen Experimente bestätigt wurde. Somit resultieren die Helligkeit und die anderen visuellen Attribute aus allen Komponentensignalen, Opponenzkanälen und melanopsinhaltigen Ganglienzellen. Durch die Entwicklung von Farbwahrnehmungsmodellen [13, 14] und Farbqualitätsmodellen [15, 16] ist die visuelle Wahrnehmung in der Licht- und Farbwissenschaft bereits relativ gut verstanden, auch wenn das nicht immer direkt und sofort in Normungen, Regelungen sowie in der Lichtproduktentwicklung berücksichtigt ist. Die visuelle Wahrnehmung und deren psychologisch-emotionalen Ausdrucksformen (wie die Raumpräferenz, die Farbharmonie und die Helligkeit) sind eher akute Lichtwirkungen, die innerhalb kurzer Zeit (im Bereich von wenigen Minuten) gebildet werden.
Vor Kurzem erfuhren die Wissenschaftler/-innen der Neurobiologie und der Lichttechnik, dass die melatoninhaltigen ipRGC sowohl für die Helligkeitswahrnehmung als auch für die Wachheit und Hormonregulation mitzuständig sind. Der Begriff image forming (IF) im Gegensatz zu nonimage forming (NIF) bezieht sich auf die Abbildung von Gegenständen auf die Netzhaut und somit auf die Wahrnehmung von Kontrasten (Hell-Dunkel-Unterschied oder Strukturunterschied). Die ipRGC ermöglichen dabei keine örtlichen und zeitlichen Kontraste (engl. nonimage forming, NIF ). Im Jahr 2019 gelang es [17], die Wirkungen und Beteiligungen von melatoninhaltigen Zellen bei niedrigen örtlichen Frequenzen bis 0,8 cpd (engl. cycle per degree , Periode pro Grad Sichtwinkel) und bis zu einer zeitlichen Frequenz bis 0,45 Hz bei einem Michelson-Kontrast von größer als 14 % nachzuweisen.
3.3.2 Verarbeitungszentren und Übertragungsbahnen für nicht visuelle Lichtwirkungen
Für die nicht visuellen Lichtwirkungen stellen die ipRGC mit ihren intrinsischen Signalbildungen in den melatoninhaltigen Pigmenten sowie mit den von den LMS-Zapfen und Stäbchen zu ipRGC eingekoppelten Signalen die Anfangsstationen dar (s. Abb. 3.10).
Die ipRGC-Signale werden zum visuellen Kortex V 1 übertragen und tragen zum visuellen Sehverhalten bei (s. [18], s. Abb. 3.9). Für die nicht visuellen Lichtwirkungen ( Abb. 3.10) werden die ipRGC-Signale sowohl zum Schrittmacher, dem SCN, als auch zu weiteren Verarbeitungszentren im Gehirn übertragen und sind für viele physiologische und psychologische Wirkungen (z. B. Wachheit, Emotionen, Lernen, Hormonbildung) mitverantwortlich.
Der SCN ( Abb. 3.10) ist eine Struktureinheit im Hypothalamus und ist etwa 0,25 mm 3pro Nukleus groß [29, 30]. Er ist der Taktgeber für den zirkadianen Biorhythmus, dessen Oszillationsfähigkeit durch zwei miteinander verbundene molekulare Regelungsschleifen ermöglicht wird. Die Zeitgeberfunktion und die Synchronisation mit der Umwelt sind mit dem SCN auf der Eingangsseite durch die ankommenden ipRGC-Signale und durch die Signale von anderen Körpereinheiten möglich. Hinzu kommen noch – auf der Ausgangsseite – die Signale zu den anderen Verarbeitungszentren im Gehirn, die Informationen über den Zeitpunkt und Phasenlage enthalten ( Abb. 3.11).
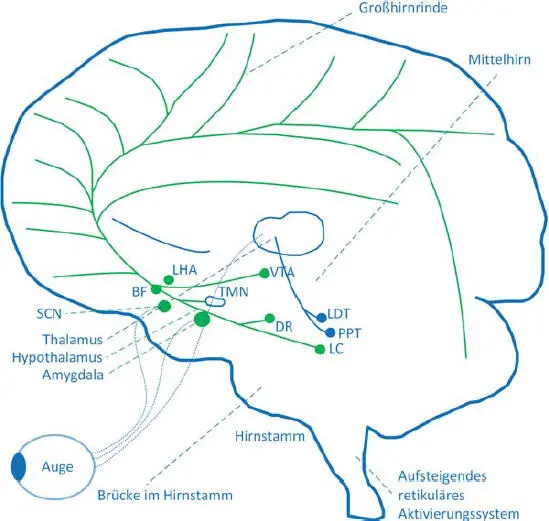
Abb. 3.10 Verarbeitungszentren und Ubertragungsbahnen für nicht visuelle Lichtwirkungen. Quelle: TU Darmstadt.
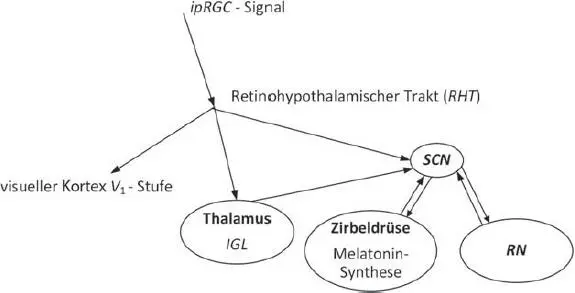
Abb. 3.11 Verarbeitungszentren und Ubertragungsbahnen des ipRGC-Signals mit dem SCN im Fokus. Quelle: TU Darmstadt.
Die Informationen von der Umwelt, die sog. photic-Informationen , gehen von den ipRGC aus und werden im retinohypothalamischen Trakt direkt zum SCN übertragen (s. Abb. 3.11). Der Signalaustausch vom SCN mit der Netzhaut kann auch indirekt über das sog. intergeniculate leaflet (IGL) im Thalamus über den sog. geniculohypothalamischen Trakt (GHT) erfolgen. Die Signale auf den beiden Bahnen RHT und GHT enthalten somit die Information über die Lichtintensität und die spektrale Zusammensetzung des einfallenden Umgebungslichts. Die Kommunikation zwischen SCN und RN (Raphe-Nuklei), eine körpereigene, sog. nonphotic communication , findet über den sog. raphe-hypothalamischen Trakt statt, der Informationen über die Phase des zirkadianen Rhythmus enthält und die Vigilanz des Körpers regelt. Der RN selbst steuert die Generation des Hormons Serotonin , ein Gewebshormon und Neurotransmitter, das sich auf das Blutsystem, Magen-Darm-System, Herz-Kreislauf-System auswirkt. Die wichtigsten Funktionen des Serotonins ist die Beeinflussung der Perzeption, der Sensorik, der Schmerzverarbeitung, des Schlafs und der Temperaturregulierung.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Beleuchtung in Innenräumen - Human Centric Integrative Lighting»
Представляем Вашему вниманию похожие книги на «Beleuchtung in Innenräumen - Human Centric Integrative Lighting» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Beleuchtung in Innenräumen - Human Centric Integrative Lighting» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.