Tran Quoc Khanh - Beleuchtung in Innenräumen - Human Centric Integrative Lighting
Здесь есть возможность читать онлайн «Tran Quoc Khanh - Beleuchtung in Innenräumen - Human Centric Integrative Lighting» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на немецком языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Beleuchtung in Innenräumen - Human Centric Integrative Lighting
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Beleuchtung in Innenräumen - Human Centric Integrative Lighting: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Beleuchtung in Innenräumen - Human Centric Integrative Lighting»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Beleuchtung in Innenräumen - Human Centric Integrative Lighting — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Beleuchtung in Innenräumen - Human Centric Integrative Lighting», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
(3.3) 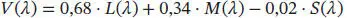
Die Gewichtung eines Spektrums optischer Strahlung mit der Hellempfindlichkeitsfunktion V (𝜆) ist die Grundlage für viele lichttechnische Bewertungsgrößen. Sie beschreibt in einer ersten (groben) Annäherung die menschliche Hellempfindung im fotopischen Bereich (Tagesehen) und ist in der Abb. 3.6zu sehen. V ′(𝜆) beschreibt die Hellempfindung im skotopischen Helligkeitsbereich (Nachtsehen). In der Abb. 3.6ist ein Beispiel für eine sog. mesopische visuelle Empfindlichkeitsfunktion (im Übergangsbereich zwischen Tagessehen und Nachtsehen) ebenfalls dargestellt (gelbe Kurve). Im mesopischen Bereich gelten – je nach Leuchtdichteniveau und Sehaufgabe – sehr unterschiedliche Empfindlichkeitsfunktionen, was wegen dessen Komplexität den Rahmen dieses Buches sprengen würde.
3.3.1.1 Horizontalzellen-Bipolarzellen-Schicht
Die Horizontalzellen der Bipolarschicht erhalten Signale von vielen Fotorezeptoren und senden diese an die Synapsen zwischen den Rezeptoren und den bipolaren Zellen. Durch ihre Fähigkeit der lateralen Signalinhibition sind sie für die Lokaladaptation oder auch für die Kontrastverstärkung im Auge wesentlich.
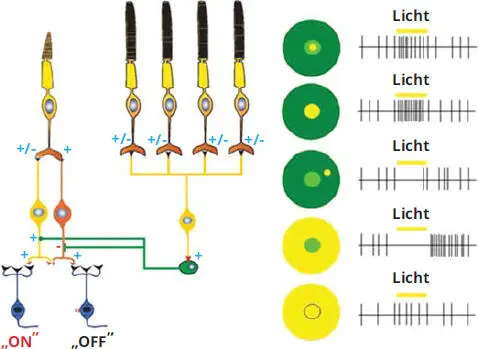
Abb. 3.7 Verschaltung in der Bipolarschicht (nach Hubel, 1989 [2]). Reproduziert mit Genehmigung von der Springer Nature Customer Service Center GmbH.
Die Bipolarzellen (s. Abb. 3.7) sind für die Reizweiterleitung von den Fotorezeptoren zu den Ganglienzellen zuständig.
Gleichzeitig wandeln die Bipolarzellen das Konstantspannungssignal der Rezeptoren in ein frequenzmoduliertes Signal um. Man unterscheidet im Wesentlichen zwei Gruppen: die ON-Bipolarzellen, die mit Stäbchen und Zapfen verschaltet sind und das negative Signal der Fotorezeptoren invertieren, und die OFF-Bipolarzellen, die nicht invertierend und nur mit den Zapfen verbunden sind. Sie dienen vor allem zum Scharfsehen in der Fovea. Weiterhin haben die OFF-Zellen keine direkte Verbindung zu den Ganglienzellen.
Die Amakrinzellen bilden eine weitere laterale Verbindung zwischen den OFF-Bipolarzellen und mehreren Synapsen zwischen den ON-Bipolarzellen und den Ganglienzellen. Dabei optimiert die Amakrinzelle die Bildung eines ON-Signals auf doppelte Weise: Sie bildet Synapsen mit ON-Bipolarzellen und gibt ihre Exzitation an die ON-Bipolarzelle weiter. Durch inhibitorische Synapsen unterdrückt die Amakrinzelle zusätzlich die Aktivität von OFF-Bipolarzellen. Durch diese Kombination aus Aktivierung von ON-Bipolarzellen und Hemmung von antagonistischen OFF-Bipolarzellen kann auch bei einer schwachen Exzitation der Stäbchen-Bipolarzelle (bei wenig Licht) schon ein brauchbares ON-Signal entstehen.
3.3.1.2 Ganglienschicht
Die Ganglienschicht besteht aus den normalen Ganglienzellen, denen – je nach Lage – ein (in der Fovea) oder mehrere Fotorezeptoren (bis zu 130 in der Peripherie) zugeordnet sind. Durch die laterale Verschaltung der Netzhautzellen hat jede Ganglienzelle ein relativ großes Einzugsgebiet. Das Gebiet, das alle auf eine individuelle Ganglienzelle wirkenden Netzhautzellen umfasst, nennt man rezeptives Feld . Man kann das Ausmaß dieser rezeptiven Felder ermitteln, indem man das Ausgangssignal einer Ganglienzelle registriert und mit einem feinen Lichtstrahl die Netzhautzellen Punkt für Punkt abtastet. Jeder Punkt, der bei Beleuchtung eineReaktion der Ganglienzelle auslöst, gehört zu dessen rezeptivem Feld.
Man findet bei solchen Untersuchungen rezeptive Felder mit variablem Durchmesser, im Mittel von 50 μm. Die Beleuchtung des inneren Bereiches (nah der Ganglienzelle, von der man ableitet) ergibt eine Aktivierung der Ganglienzelle (ein sog. ON-Response ). Außerhalb dieses zentralen ON-Feldes erzeugt man durch Belichtung eine Hemmung der Ganglienzelle, denn dabei überwiegt die laterale Inhibition die Hemmung der Ganglienzelle durch ihre Nachbarzellen.
Die Erkenntnisse über die Verschaltung der normalen Ganglienzellen und der intrinsisch fotoempfindlichen Ganglienzellen (ipRGC) miteinander und zu den Rezeptoren sowie daraus die Beiträge der Zapfen- und Stäbchensignale zu visuellen und nicht visuellen Verarbeitungen sind in den letzten Jahren durch intensive Forschungsarbeiten der Neurobiologie und Zellenforschung stark angewachsen. In der Abb. 2.2sind die Verschaltungen der Signale der LMS-Zapfen und der Stäbchen zueinander und zu den Bipolarzellen veranschaulicht, die wiederum miteinander über die Amakrinzellen kommunizieren und Signale an die ipRGC übertragen. Die ipRGC sind selbst fotoempfindlich und absorbieren Photonen in deren melanopsinhaltigen Pigmenten. Das gesamte Signal einer ipRGC besteht somit aus den Signalen aus den Zapfen, Stäbchen und aus den selbst generierten Signalen. Die normalen Ganglienzellen enthalten dagegen nur Signale aus den Zapfen und den Stäbchen.
Eines der wichtigsten Ergebnisse der jüngsten neurobiologischen Forschung ist in [3] beschrieben. Ein neuer Typ von Amakrinzellen wurde identifiziert, die auf der Eingangsseite anregende Eingangssignale von ON-Bipolarzellen erhalten, die mit S-Zapfen direkt verbunden sind. Auf der Ausgangsseite dieser Amakrinzellen werden hemmende Signale an die ipRGC übertragen. Diese Ergebnisse sind insofern bedeutungsvoll, da sie zeigen, dass die Signale der ipRGC immer eine Komponente von S-Zapfen enthalten, die eine signalhemmende Wirkung aufweisen. Eine Vergrößerung der Signale der S-Zapfen führt zu einer Reduktion der ipRGC-Signale und zu einer Vergrößerung des Augenpupillendurchmessers [4, 5].
3.3.1.3 Die Sehbahn
Die Verarbeitung der visuellen Information verläuft über die verschiedenen Systeme der Sehbahn voneinander getrennt. Im sog. P-System (parvozellulär) werden Informationen zur Form und Farbe von Objekten transportiert und verarbeitet. Im sog. M-System (magnozellulär) werden dagegen Orts- und Bewegungsinformationen verarbeitet und weitergeleitet. In Abb. 3.8ist der Verlauf der Sehbahn dargestellt.
Chiasma opticum : In der Sehbahnkreuzung werden die Gesichtsfelder sortiert (s. Abb. 3.8). Die jeweils rechten Sehfelder beider Augen werden in die rechte Gehirnhälfte geleitet, die linken in die linke. Oberhalb des Chiasmas sitzt der suprachiasmatische Nukleus (SCN). Dieses Kerngebiet verarbeitet einen kleinen Teil der nicht visuellen Augeninformation, er bildet eine Komponente der inneren Uhr und ist für zirkadiane Rhythmen zuständig, worüber später in diesem Buch berichtet wird. Speziell werden hier auch die sog. Sakkaden gesteuert. Sakkaden sind kurze, ruckartige Bewegungen der Augen, die unbewusst ablaufen und das Gesichtsfeld abtasten.
Seitliche Kniehöcker: Die seitlichen Kniehöcker sind Teile des Thalamus, der den größten Teil des Zwischenhirns bildet und sich aus vielen eng vernetzten Kerngebieten zusammensetzt. Hier laufen alle Signale von Sinnesorganen zusammen und werden an die Gehirnrinde weitergegeben. Die Bezeichnung „ Tor zum Bewusstsein “ weist auf die Leitfunktion des Thalamus für sensorische Information hin. Der Thalamus kann verschiedene Signale regulieren, um „wichtigeren“ Eindrücken Vorrang zu geben. Im seitlichen Kniehöcker werden die visuellen Eindrücke gefiltert.
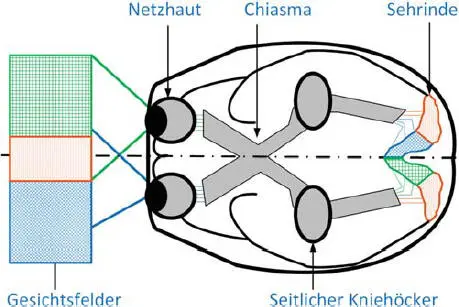
Abb. 3.8 Sehbahn im Gehirn. Quelle: TU Darmstadt [1].
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Beleuchtung in Innenräumen - Human Centric Integrative Lighting»
Представляем Вашему вниманию похожие книги на «Beleuchtung in Innenräumen - Human Centric Integrative Lighting» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Beleuchtung in Innenräumen - Human Centric Integrative Lighting» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.