Tran Quoc Khanh - Beleuchtung in Innenräumen - Human Centric Integrative Lighting
Здесь есть возможность читать онлайн «Tran Quoc Khanh - Beleuchtung in Innenräumen - Human Centric Integrative Lighting» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на немецком языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Beleuchtung in Innenräumen - Human Centric Integrative Lighting
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Beleuchtung in Innenräumen - Human Centric Integrative Lighting: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Beleuchtung in Innenräumen - Human Centric Integrative Lighting»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Beleuchtung in Innenräumen - Human Centric Integrative Lighting — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Beleuchtung in Innenräumen - Human Centric Integrative Lighting», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Im Abschn. 9.4.2dieses Buches werden Lichtwirkungen in den nächtlichen Stunden mit polychromatischem weißem Licht, u. a. die Phasenverschiebung in der Abb. 9.31 und in der Tab. 9.5, ausführlich beschrieben. In [32] wird über eine Forschungsarbeit berichtet, die die Wellenlängenabhängigkeit der Phasenverschiebung des Melatoninrhythmus untersuchte. Die untersuchten Wellenlängen waren dabei 660 nm (rot), 595 nm (bernsteinfarben, engl. amber ), 525nm (grün), 497nm (grünblau) und 470nm (blau). Die Lichtquellen waren Halbleiter-LEDs. Fünfzehn Testpersonen nahmen am Test mit allen Wellenlängen sowie an einer Referenzbedingung mit keinem Licht teil. Jede Lichtbedingung durchlief zwei aufeinander folgende Nächte (Nacht 1 und Nacht 2). Die Speichelproben für die Melatoninanalyse wurden immer jeweils 30 min von 19:00–2:00 Uhr (Nacht 1) und von 19:00– 1:00 Uhr (Nacht 2) entnommen. Die Lichtexposition bei jeder Testbedingung wurde für die Nacht 1 zwischen Mitternacht und 2:00 Uhr durchgeführt. Der Zeitpunkt der Melatoninanstiege (engl. dim light melatonin onset , DLMO ) in den beiden Nächten wurde jeweils gemessen, woraus die Phasenverschiebung errechnet wurde. Die kürzeren Wellenlängen (470 und 497 nm) verursachten die größere Phasenverschiebung zwischen 27 und 36 min.
3.4.4 Chronotypen, Schlafverhalten
Die Bestimmung der freilaufenden zirkadianen Periodendauer kann anhand der Bestimmung der Periodizität der Verläufe von Melatoninausschüttung, Cortisolsynthese und Körperkerntemperatur (CBT) realisiert werden. Eine Forschungsarbeit dazu wurde an zwei Altersgruppen von Czeisler et al. [33] durchgeführt, deren Ergebnisse in der Tab. 3.1dargestellt sind.
Aus der Tab. 3.1geht hervor, dass die intrinsische zirkadiane Periodendauer nicht vom Alter und vom physiologischem Mechanismus (CBT, Cortisol, Melatonin) abhängt. Diese Mechanismen unterliegen also der zentralen periodischen Steuerung des zentralen Schrittmachers.
Tab. 3.1 Freilaufende intrinsische zirkadiane Periodendauer. Quelle: Basierend auf Czeisler et al., 1999 [33].
| Altersgruppe | Mittleres Alter (Jahre) | Periodendauer CBT (h:min) | Periodendauer Melatonin (h:min) | Periodendauer Cortisol (h:min) |
|---|---|---|---|---|
| Jung | 23,7 ± 2,7 | 24:10 | 24:11 | 24:11 |
| Älter | 67,4 ± 3,2 | 24.10 | 24:13 | Keine Angabe |
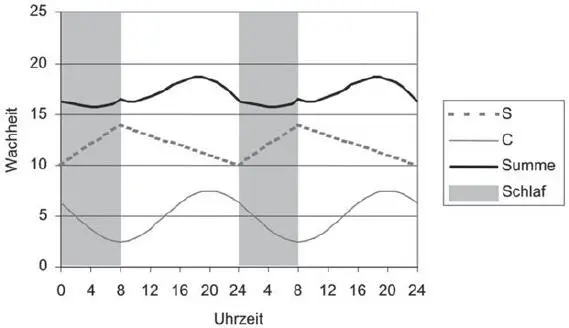
Abb. 3.15 Zirkadiane und homöostatische Komponenten der Wachheit über zwei Tage. Quelle: Basierend auf Vandahl et al., 2009 [20]/Technische Universität Ilmenau.
Die freilaufende zirkadiane Periodendauer verteilt sich innerhalb der Bevölkerung statistisch gesehen normal . Diejenigen, die eine kürzere Periodendauer haben, sind die sog. Lerchen , stehen morgens früh auf, sind abends früh müde und möchten deshalb früh ins Bett. Diejenigen Menschen, die eine längere Periodendauer haben (sog. Eulen ), verhalten sich umgekehrt. Sie stehen später auf und sind in den Abendstundenwie auch in den ersten Nachtstunden munter. Mit einem durchschnittlichen Wert der freilaufenden zirkadianen Periodendauer von etwa 24 h und 12 min tickt die biologische Uhr der meisten Menschen etwas zu langsam und muss deshalb durch das tägliche Morgenlicht vorgestellt werden.
Der Wach-Schlaf-Zyklus enthält Verläufe von Wachheit am Tag und Schlafphasen in der Nacht und setzt sich in einer ersten Näherung additiv aus einem zirkadianen Rhythmus mit einer zirkadianen Komponente C (s. Abb. 3.15) und aus einer homöostatischen Komponente S des sog. Schlafhomöostaten zusammen. Beide Komponenten verlaufen über den Tag periodisch.
Von etwa 20–8 Uhr sinkt die Wachheit in der zirkadianen Komponente (s. Abb. 3.15) und erreicht ein Minimum um 8 Uhr. Danach erhöht sich die Wachheit stetig bis ca. 20 Uhr. Während eines angenommenen Schlafs zwischen 0–8 Uhr sinkt diese zirkadiane Komponente, während der homöostatische Schlafdruck sinkt und die homöostatische Wachheit ansteigt. In der Summe sinkt die Wachheit während des nächtlichen Schlafs bis etwa 5 Uhr am frühen Morgen und steigt danach an. Am Tag zwischen etwa 8–20 Uhr sinkt die homöostatische Wachheit wieder stetig, während die zirkadiane Komponente ansteigt. Als Ergebnis erhöht sich die Summe der Wachheit am Tag zwischen etwa 5–18 Uhr, um danach wieder zu sinken. Man kann auch feststellen, dass die beiden Komponenten zwischen 0– 20 Uhr gegenläufig verlaufen. Sie verlaufen in der gleichen Richtung zwischen 20 Uhr und Mitternacht (0 Uhr) und helfen den Menschen, sich auf den Schlaf vorzubereiten.
Die Lichtwirkungen in den nächtlichen Stunden sind in Abschn. 9.4.2beschrieben. Die Wachheit in den Tagesstunden ist in Abschn. 9.7.2detailliert dargestellt.
3.5 Ausgangsgrößen des visuellen und des nicht visuellen Gehirnverarbeitungsapparats: eine Systematik
In den vorangegangenen Abschnitten wurden die Eingangsgrößen des Gesamtsystems der Gehirnverarbeitung sowie dessen visuelle und nicht visuelle Verarbeitungsnetzwerke dargestellt. In diesem vorliegenden Abschnitt werden die Ausgänge dieses Systems beschrieben. Diese Ausgangsgrößen werden in physiologisch-fotometrische Kenngrößen (Sehleistung/Arbeitsleistung), in farbmetrisch-psychologisch-emotionale Attribute und in nicht visuelle Metriken unterteilt. Es ist allerdings nicht immer möglich, diese drei Gruppen so klar voneinander zu trennen. Die Emotionen haben z. B. zum guten Teil mit der Wachheit zu tun und eine kurze Reaktionszeit hat ebenfalls ihre Wurzeln in einem hohen Helligkeitskontrast. Die Separation der Metriken dient dem Zweck, die hohe Komplexität der Signalverarbeitung in einzelne Teilthemen zu zerlegen und auch in diesem Buch diese Teilthemen in einzelnen Kapiteln ergebnisorientiert zu beschreiben. Das gesamte Ergebnis der Darstellungen in den einzelnen Kapiteln mündet in eine gesamte Beleuchtungskonzeption in Kap. 12am Ende dieses Buches.
Die Systematik der Ausgangsgrößen ist in der Abb. 3.16zusammenfassend dargestellt.
Aus der Abb. 3.16geht Folgendes hervor:
• Die fotometrischen Kenngrößen dienen hauptsächlich der Charakterisierung der Sehleistung und der Arbeitsleistung, wie sie in Kap. 4dargestellt werden.
• Die Helligkeit und visuelle Klarheit, die die bisherigen, rein fotometrischen Kenngrößen mit den Signalen der ipRGC und der S-Zapfen beinhalten, werden in Kap. 5vorgestellt.
• Die Farbqualität bildet die Schwerpunkte der Licht- und Farbforschung seit etwa 2002 bis heute. Die Forschungsarbeiten erzielten seitdem große Fortschritte und werden in Kap. 6diskutiert.
• Die beiden Komponenten Farbqualität und Arbeitsleistung/Sehleistung führen zu der neuen Modellierung und Verifikation der Lichtqualität in Kap. 7.
• Der visuelle Komfort, die Leuchtdichteverteilung der Wände und der Decke, die stroboskopischen Effekte bilden weitere Aspekte der Beleuchtungsqualität in Kap. 9.
• Die nicht visuellen Aspekte werden in diesem vorliegenden Kap. 3und danach auch ausführlich in Kap. 9erörtert.
• Alle o. g. Aspekte führen zu einem Konzept der Beleuchtungsplanung bzw. Lichtplanung in Kap. 12. Das ist das Ergebnis der Strategie und Philosophie unter dem Oberbegriff Human Centric Lighting oder Integrative Lighting. Diese beiden Begriffe haben unterschiedliche Historien aber den gleichen Inhalt und die gleichen Ziele.
3.6 Grundaspekte von Human Centric Lighting – Integrative Lighting [35]
Интервал:
Закладка:
Похожие книги на «Beleuchtung in Innenräumen - Human Centric Integrative Lighting»
Представляем Вашему вниманию похожие книги на «Beleuchtung in Innenräumen - Human Centric Integrative Lighting» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Beleuchtung in Innenräumen - Human Centric Integrative Lighting» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.