Dr. Freia Hünig - Das Psoriasis-Syndrom
Здесь есть возможность читать онлайн «Dr. Freia Hünig - Das Psoriasis-Syndrom» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на немецком языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.
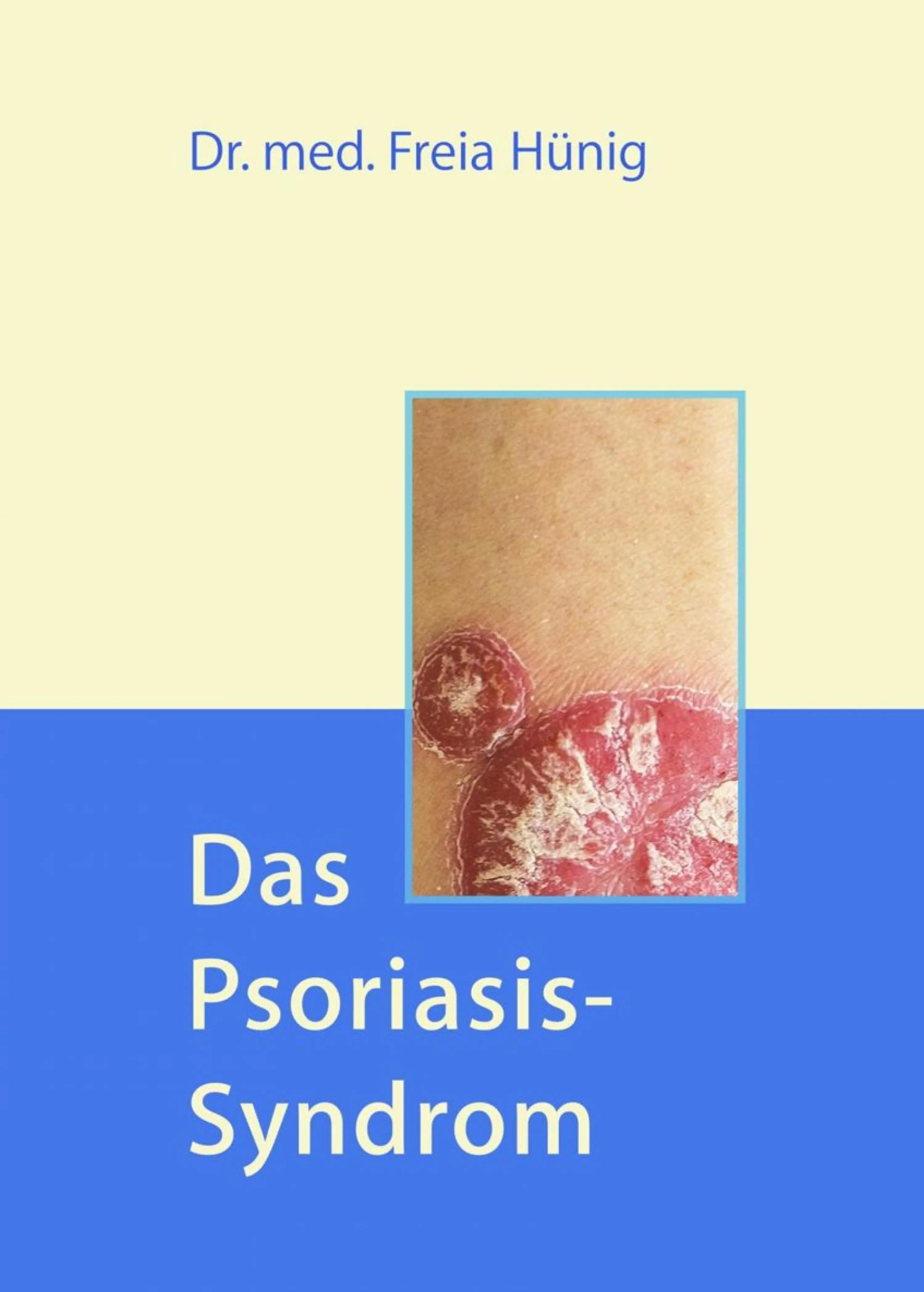
- Название:Das Psoriasis-Syndrom
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Das Psoriasis-Syndrom: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Das Psoriasis-Syndrom»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Die Arbeit konkretisiert die zentrale Bedeutung von cAMP für die zellulären Calciumsignale, die calciumabhängigen Differenzierungsprozesse, den Aufbau des epidermalen Calciumgradienten und den hier vorgestellten epidermalen Calciumkreislauf. Es wird dargestellt, wie sich aus einer fatal blockierten cAMP-Versorgung unter Aktivierung der Hedgehog-Transkription die psoriatische Läsion entwickelt, und in wiefern bereits die präsymptomatische psoriatische Haut u.a. durch eine vermehrte Expression NF-κB-abhängiger Zellprodukte vom Gesunden abweicht.
Konkrete Forschungsvorschläge sollen die Entwicklung neuer, ergänzender Therapien anregen.
Das Psoriasis-Syndrom — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Das Psoriasis-Syndrom», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
3 Die Ergebnisse von G. Hong et al. (2010) weisen in dieselbe Richtung: ihren Forschungen entsprechend wird die IL-1ß/IFNγ induzierte Aktivierung von NF-κB in Hepatozyten durch inhibitorische Effekte von cAMP auf IKK und die IκB-Phosphorylierung verhindert. Diese cAMP-Effekte sind jedoch PKA unabhängig [224]. A. Kim et al. (2009) haben egänzend gezeigt, dass auch in Melanomzellen ein erhöhter cAMP-Spiegel die Aktivierung von NF-κB inhibiert [220].
Wie für Monozyten, T-Zellen, Astrozyten, Hepatozyten und Melanomzellen nachgewiesen, hebt ß2AR-Stimulation bzw. ein erhöhter zellulärer cAMP-Spiegel das zelluläre IκB- bzw. IκBα -Level an. Obwohl zurzeit noch keine expliziten Informationen über den Einfluss von cAMP auf den IκB-Gehalt von KC zu erhalten sind, kann folgendes festgehalten werden: steigende zytosolische cAMP-Spiegel unterdrücken NF-κB induzierter Genexpression (s.o.), und vermitteln so die antiinflammatorischen Wirkungen der ß2AR (s. 4.12 , 4.14.1 ), [225]. Bei einem cAMP-Mangel fehlt nicht nur die PKA/CREB-abhängige Blockade der Transkriptionsaktivität von NF-κB, sondern es fallen auch die das IκB-Level anhebenden Wirkungen von cAMP flach. Daher ergibt sich folgende, für o.g. Zelltypen gesicherte, Aussage:
Ein hoher zytosolischer cAMP-Gehalt blockiert NF-κB abhängige Genexpression, hebt das zelluläre IκB-Level und vermindert aktives NF-κB. Umgekehrt führt ein zytosolischer cAMP-Mangel zur Enthemmung von NF-κB und zu einer vermehrten Expression NF-κB-abhängiger Zellprodukte.
3.2.7 Escape-Mechanismen der ß2AR
Bei chronischer Überstimulierung von ßAR, wie z.B. unter einer ß-Mimetika-Therapie bei Asthma bronchiale oder COPD, können ß2AR (aber nicht ß1AR) an hemmende Gi-Proteine koppeln, und so zur Reduktion von cAMP und zur Schwächung katecholaminerger Wirkungen führen [226]. ß2AR können unter bestimmten Ausnahmebedingungen auch an Gq-Proteine koppeln [227].
3.3 α-Adrenozeptoren (αAR):
αAR sind wie ßAR membranständige GPCR und kommen in den Subtypen α1 und α2 vor. α1AR sind besonders im Zentralnervensystem und an der glatten Muskulatur von Gefäßen und im Urogenitaltrakt vertreten, wo sie über die Kontraktion glatter Muskulatur zur Blutdruckerhöhung und ggf. zum Harnverhalt führen, während α2AR sich im zentralen und peripheren Nervensystem finden und dort präsynaptisch die Freisetzung von Neurotransmittern hemmen.
3.3.1 Signalübertragung der α1AR
α1AR koppeln an Gq-Proteine, die ihre Impulse über Phospholipase C (PLC), Inositoltrisphosphat (IP3), Diacylglycerin (DAG), Ca2+ und Phosphokinase C (PKC) weiterleiten, wobei die PLC in einem ersten Schritt zur Bildung von IP3 und DAG aus membrangebundenem Phosphatidylinositolbisphosphat (PIP2) führt. IP3 bindet darauf an seinen Rezeptor (IP3R), der sich auf den Membranen intrazellulärer Ca2+-Speicher wie ER und SR befindet und löst dort eine Ca2+-Freisetzung in das Zytosol aus. DAG und Ca2+ aktivieren zusammen die PKC, die ihrerseits zahlreiche Stoffwechselleistungen durch Phosphorylierung ermöglicht oder unterbindet. U.a. inhibiert die PKC die AC9 und damit die cAMP-Synthese [182].
3.3.2 Calcium-Regulation durch Gq-Protein gekoppelte Rezeptoren
Gq-Protein Aktivierung ist regelmäßig von einer Ca2+-Mobilisierung aus intrazellulären Speichern wie ER oder SR begleitet. Dabei kommt es, abhängig von dem Füllungszustand der Calciumspeicher, zur Aktivierung von Zellmembran ständigen “store-operated calcium channels“ (SOCC) [228], die einen Ca2+-Auffüllungsstrom aus dem Extrazellulärraum zur Regeneration der intrazellulären Ca2+-Speicher auslösen [229]. Zusätzlich zu diesem „Store operated Ca2+-Entry“ (SOCE) können Gq-gekoppelte Rezeptoren über Aktivierung der PLC zu einem „Receptor operated Ca2+-Entry“ (ROCE) führen. Während der zytosolischen Ca2+-Erhöhung lagern sich Ca2+-Ionen an intrazelluläre Ca2+-bindende Proteine wie z.B. Calcineurin oder Calmodulin (CaM) an und aktivieren diese. Aktiviertes CaM kann u.a. ACs (außer der AC9), die lösliche Guanylatzyklase (sGC), Proteinkinasen und auch die endotheliale NO-Synthase (eNOS) aktivieren [230].
3.3.3 Signalübertragung der α2AR
α2AR koppeln an G i/0-Proteine, die eine Hemmung von Adenylatzyklasen bewirken und damit zu einer Senkung des cAMP-Spiegels und einer Blockade cAMP-abhängiger Wirkungen führen.
3.4 Cholinerge Rezeptoren (AChR)
Cholinerge Rezeptoren sind membranständige, durch ACh erregbare Rezeptoren, die als nikotinische (nAChR) oder muskarinische (mAChR) Rezeptoren vorliegen.
3.4.1 Nikotinische Rezeptoren
Nikotinische Rezeptoren (nAChR) finden sich insbesondere auf der neuromuskulären Endplatte, wo sie zu einer extrem schnellen Signalübertragung vom motorischen Nerven auf die angesteuerte Muskulatur führen. nAChR finden sich u.a. auch in den vegetativen Ganglien auf der Postsynapse des zweiten Neurons, auf KC ( 3.1), Immunzellen [231], EC [232] und, wie Kurzen (2004) zeigen konnte, auf den Schweiß produzierenden Azinuszellen der Schweißdrüsen [233: S. 455]. Die nAChR gehören zu den Liganden gesteuerten Ionenkanälen, sind also Rezeptor und Ionenkanal in einem. Ihre Kanalröhrchen werden aus fünf Einheiten (Pentamere) gebildet, welche jeweils Kombinationen aus gleichen oder verschiedenen Untereinheiten sind [234: S. 6]. Die Untereinheiten haben unterschiedliche Funktion und kommen als α-, ß-, γ- und δ-Formation vor, wobei die α-Formation, die in den Typen α1 bis α10 vorliegt, die ACh-Bindung des Rezeptors vermittelt. Jedes Pentamer muss mindestens zwei α-Untereinheiten besitzen, um ACh binden zu können. Die Aktivierung der nAChR führt über eine Konformationsänderung zur vermehrten Durchlässigkeit des Rezeptors für bestimmte Kationen (Na+, Ca2+, K+). Homopentamere werden aus fünf α7- oder α9-Einheiten gebildet [233] und sind hauptsächlich für Ca2+ durchlässig [235], während Heteropentamere aus verschiedenen Einheiten gebildet werden und unterschiedliche Leitfähigkeiten besitzen. nAChR führen durch Kationen-Einstrom unmittelbar zu einer Depolarisation der Zellmembran, der eine Aktivierung spannungsabhängiger Ca2+Kanäle (VOCs) mit forciertem Calciumioneneinstrom folgen kann [235]. In der gesunden Epidermis werden im Stratum basale α9-nAChR und im Stratum granulosum α7-nAChR exprimiert [233: S. 454].
Besonderheit: α9-nAChR sind durch ACh und Cholin, α7-nAChR durch ACh, Cholin und Muskarin stimulierbar [233: S. 456, 236, 237]. Atropin, besonders als Blocker muskarinischer Rezeptoren bekannt, blockiert cholinerge Rezeptoren mit folgender, absteigender Effektivität: mAChR > α9-nAChR > α7-nAChR > α3ß-nAChR [238].
Die nAChR kommen in drei Funktionszuständen vor: erregt&geöffnet, geschlossen&desensitisiert, geschlossen&erregbar. Diese Funktionszustände werden nacheinander durchlaufen, wobei Phosphorylierungen am Rezeptor zu einer beschleunigten Desensitisierung führen [234: S. 9-10]. Die Sensitivität von nAChR wird u.a. durch cAMP/PKA-Aktivität reguliert [239: S. 430, 240: S. 6 unten].
3.4.2 Muskarinische Rezeptoren
Muskarinische Rezeptoren (mAChR) sind GPCR, kommen in den Subtypen M1-M5 vor und werden in weiten Bereichen des Körpers direkt von parasympathischen Neuronen angesteuert. M1 ist der neuronale Typ und tritt besonders im Gehirn und ganglionär auf, M2 findet sich am Herzen und vermittelt dort die Vaguseffekte, M3 ist besonders auf glatter Muskulatur, Endothel, Drüsenzellen, Schweißdrüsen und den Belegzellen des Magens lokalisiert und führt u.a. zur Vasodilatation durch NO-Bildung oder zur Salzsäureproduktion im Magen. Über M4 und M5 ist bekannt, dass sie hauptsächlich im ZNS vorkommen; aber ihre physiologische Bedeutung ist noch weitgehend unklar [241: S. 572]. KC können alle Subtypen der M1-M5 mAChR exprimieren [159: S. 169, 242], wobei basale KC der gesunden Epidermis besonders M2, M3 und M4 und suprabasale M1 und M4 exprimieren [159, 160, 243]. Eine Aktivierung von mAChR der KC kann Einfluss auf deren Migrationsgeschwindigkeit, Apoptose, Zell-Zellkontakte und Proliferation nehmen [157].
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Das Psoriasis-Syndrom»
Представляем Вашему вниманию похожие книги на «Das Psoriasis-Syndrom» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Das Psoriasis-Syndrom» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.