Markus Kipp - Neuroanatomie
Здесь есть возможность читать онлайн «Markus Kipp - Neuroanatomie» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на немецком языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.
- Название:Neuroanatomie
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Neuroanatomie: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Neuroanatomie»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Neuroanatomie erklärt anschaulich die wichtigsten und prüfungsrelevanten funktionellen und anatomischen Zusammenhänge des Zentralen Nervensystems. Das Buch eignet sich daher ideal zum Nachschlagen, Lernen und Verstehen in jeder Lernphase: für die Kurs- und Semesterbegleitung, fürs nächste Testat, zur effektiven Vorbereitung auf die Semesterklausur oder das Physikum. Hinzu kommt als neues Konzept die didaktische Verzahnung von schematischen Grafiken und detaillierten Fotografien von Gehirn- und Rückenmarkspräparaten. Damit werden Theorie und Praxis präzise und anschaulich miteinander verbunden und die Lerneffekte intensiviert. Die inhaltliche Gliederung sorgt für Struktur und prompten Zugriff auf den Lernstoff: • Zunächst werden die Grundlagen der Neuroanatomie vorgestellt. • Die einzelnen Bestandteile und Strukturen des Zentralen Nervensystems werden anschließend im Hauptteil erklärt. • Es folgt eine Betrachtung ihrer Zusammenhänge und ihres ineinandergreifenden Zusammenspiels. • Ein eigenes Kapitel ist den bildgebenden Verfahren und ihrer Anwendung in der Neuroanatomie gewidmet. Das Buch ist abwechslungsreich und didaktisch zielführend konzipiert: Ein übersichtliches, farbkodiertes Layout und durchdacht angeordnete Legenden ermöglichen einen raschen Überblick über den Lernstoff. Die komplexen Zusammenhänge der Neuroanatomie lassen sich mit diesem Konzept daher von Grund auf leicht verständlich erschließen und einprägen. Darüber hinaus enthält das Buch praktische Merkboxen und Hervorhebungen für spannende klinische, pharmakologische und wissenschaftliche Themen. Am Ende eines jeden Kapitels wird noch einmal gesondert auf die wichtigsten IMPP-relevanten Besonderheiten eingegangen. So kann das Gelernte auch immer an den MC-Fragen direkt getestet werden.
Neuroanatomie — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Neuroanatomie», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Oligodendrozyten und Schwann-Zellen
Oligodendrozyten und Schwann-Zellen bilden Myelin. Es handelt sich hierbei um eine lipidreiche Biomembran, welche die Axone der meisten Nervenzellen von Wirbeltieren spiralförmig umgibt und somit elektrisch isoliert. Myelin wurde 1854 von dem Pathologen Rudolf Virchow (1821–1902) mittels Lichtmikroskopie an Gewebeschnitten entdeckt. Er fand in Nervenfasern eine Markscheide und schlug vor, sie Myelin(griech. μυελός - „Mark“) zu nennen ( Abb. 1.12).
Abb. 1.12
Myelinscheiden
Elektronenmikroskopische Aufnahme eines myeliniserten Axons im Bereich des Corpus callosum. Die Myelinscheide stellt sich in dunkelgrauen Schichten dar, die um das hellere Axon gewickelt sind.
Im Vergleich zu anderen Biomembranen weist Myelin einen besonders hohen Lipidgehalt (70 %) und einen relativ geringen Proteinanteil (30 %) auf. Daher erscheint Myelin in der makroskopischen Sicht weiß, weshalb stark myelinisierte Regionen im Zentralnervensystem auch als „weiße Substanz“ bezeichnet werden, im Gegensatz zur gering myelinisierten „grauen Substanz“. Darauf werden wir im nächsten Kapitel noch genauer eingehen. Myelinscheiden findet man nicht nur um die Axone des zentralen sondern auch des peripheren Nervensystems. Sowohl im zentralen als auch peripheren Teil des Nervensystems sind die Myelinscheiden entlang der Axone regelmäßig von den Ranvier-Schnürringenunterbrochen (siehe Lehrbücher der Physiologie). Nur an den Ranvier-Schnürringen entstehen Aktionspotenziale, nicht aber in den myelinisierten Bereichen des Axons (Internodien). Dieser Aufbau ermöglicht die saltatorische Erregungsleitung, welche deutlich schneller als die kontinuierliche Erregungsleitung nicht-myelinisierter Fasern ist. Außerdem spart diese Art der Erregungsleitung Energie, da ein Aktionspotenzial nur am Ort der Schnürringe und nicht kontinuierlich entlang eines Axons aufgebaut werden muss. Myelin wird im Zentralnervensystem von Oligodendrozyten, im peripheren Nervensystem von Schwann-Zellengebildet. Ein wichtiger Unterschied bei den myelinbildenden Zellen besteht darin, dass eine Oligodendrogliazelle mehrere Axone mit Myelin versorgt, während jede Schwann- Zelle nur ein einziges Axon mit Myelin umgibt ( Abb. 1.13).
Abb. 1.13
Myelinisierung von Axonen
In der Peripherie werden die Myelinscheiden von Schwann-Zellen produziert. Im Zentralnervensystem übernimmt diese Funktion der Oligodendrozyt.
Durch die Myelinscheide springt ein Aktionspotenzial von einem Ranvier-Schnürring zum nächsten.
Die schnelle Weiterleitung des Aktionspotenzials ist funktionell von großer Bedeutung. Um die Geschwindigkeit dieses Impulses zu erhöhen, hat die Evolution zwei unabhängige Mechanismen entwickelt. Ein Mechanismus besteht darin, den Axondurchmesser zu vergrößern, wie es z. B. beim Tintenfisch der Fall ist (hier gilt: je größer der Durchmesser eines Axons, desto schneller die Leitungsgeschwindigkeit). Er beträgt hier fast einen Millimeter! Der zweite Mechanismus ist die Myelinisierung, also das Umwickeln des Axons mit den Membranen von Oligodendrozyten oder Schwann-Zellen. Funktionell wird durch die Myelinisierung die Membrandicke des Axons erheblich vergrößert und die Leitungsgeschwindigkeit stark erhöht.
Forschung
Der Durchmesser des Axons und die Dicke der Myelinschicht stehen in einem direkten Zusammenhang. Je dicker ein Axon, desto dicker auch seine Myelinschicht und desto schneller seine Leitungsgeschwindigkeit. Dieser Umstand kann in der Forschung ausgenutzt werden. Axone, die im Rahmen einer demyelinisierenden Erkrankung, wie etwa der Multiplen Sklerose, ihre Myelinschicht verlieren, können sich regenerieren: man spricht von Remyelinisierung. Dieser Regenerations-Mechanismus ist jedoch nicht ganz so effektiv wie die Myelinisierung im Rahmen der Entwicklung. Die Folge ist, dass die Dicke der Myelinschicht im Verhältnis zur Dicke des Axons dünner als gewöhnlich ausgebildet ist. So kann erkannt werden, ob eine Remyelinisierung stattgefunden hat oder nicht.
Saltatorische Erregungsleitung
Die Reizweiterleitung durch elektrische Impulse ist eine Gemeinsamkeit, die alle Lebewesen miteinander teilen. Dennoch gibt es z. B. bei der Erregungsweiterleitung Unterschiede:
Bei einer kontinuierlichen Erregungsleitung wird die Erregung durch das Axon mittels einer fortlaufenden Bildung des Aktionspotenzials weitergeleitet. Folglich muss an jeder Stelle des Axons eine Depolarisation stattfinden. Eine kontinuierliche Erregungsleitung ist vor allem bei wirbellosen Tieren wie Tintenfischen oder Regenwürmern die Form der Erregungsweiterleitung. Tintenfische besitzen besonders dicke Axone (Riesenaxon), zurückzuführen auf evolutionäre Gründe: Die Geschwindigkeit der Erregungsleitung lässt sich bei der fortlaufenden Bildung eines Aktionspotenzials nur durch eine Vergrößerung des Durchmessers steigern. In Folge dessen sinkt der Innenwiderstand des Axons und das Aktionspotenzial kann schneller gebildet werden.
Wirbeltiere besitzen im Gegensatz zu den eben genannten Tintenfischen nach außen hin eine Isolierung des Axons. Fettreiche Lipide und Eiweiße bilden die sogenannte Myelinscheiden und umhüllen fortlaufend das Axon. Diese sind lediglich an den Ranvier-Schnürringen unterbrochen. Auf diese Weise kann ein Aktionspotenzial nur an den nicht isolierten Ranvier-Schnürringen gebildet werden. Im Vergleich zur kontinuierlichen Erregungsleitung läuft die saltatorische um ein Vielfaches schneller ab. Die Erregung ‚springt‘ innerhalb des Axons von Ranvier-Schnürring zu Ranvier-Schnürring und überbrückt die nach außen hin isolierenden Myelinscheiden. Eine Depolarisation kann nur an den unisolierten Schnürringen erfolgen. Bei der kontinuierlichen Erregungsleitung gibt es keine Myelinscheiden. Die Axone müssen fortlaufend depolarisiert werden, was mehr Zeit und Energie in Anspruch nimmt.
| Kontinuierliche Erregungsleitung | Saltatorische Erregungsleitung | |
| Anzutreffen bei: | Wirbellosen | Wirbeltieren |
| Geschwindigkeit: | bis zu 30 m/s | bis zu 100 m/s |
| Größe derAxons: | bis zu 2 mm | vom Durchmesser deutlich dünner |
| Axonale Isolierung: | lediglich die natürliche Isolierung des Axons (wenig wirkungsvoll) | lipidreiche Myelinscheiden isolieren das Axon |
| Ort der Depolarisation: | fortlaufend am gesamten Axon | nur an den Ranvier-Schnürringen |
Mikrogliazellen
Als Mikroglia oder Mesoglia bezeichnet man eine Gruppe von Immuneffektorzellen des Zentralnervensystems. Sie werden zwar formal zur Familie der Gliazellen gerechnet, im eigentlichen Sinn handelt es sich jedoch um Zellen des mononukleär-phagozytären Systems. Es wird davon ausgegangen, dass sie im Laufe der Entwicklung in das Zentralnervensystem einwandern (ganz ähnlich wie etwa die Kupffer-Zellen der Leber). 9 , 10 Wie aus ihrem Namen bereits abgeleitet werden kann, handelt es sich bei der Mikroglia um die kleinste Gliazellpopulation. Mikroskopisch sieht man schmale, lang gestreckte Zellen, die einen irregulären, länglichen Zellkern mit dichtem Chromatin besitzen. Die Zellfortsätze können fein und sehr verzweigt sein (ramifizierte, ruhende Mikroglia; Abb. 1.14).
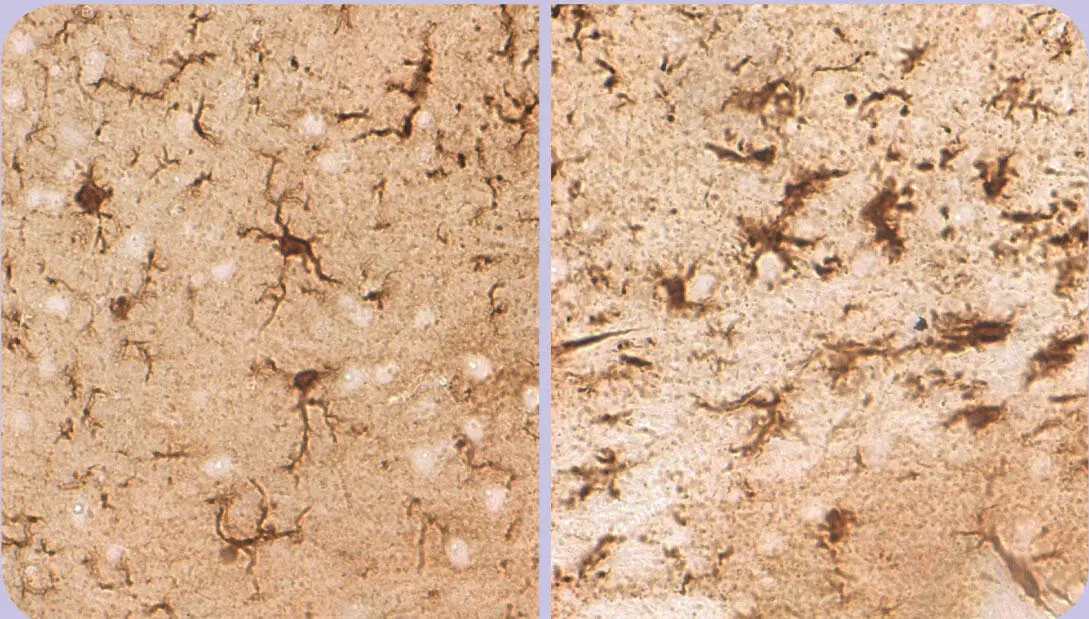
Abb. 1.14
Ruhende (li) und aktivierte (re) Mikroglia
Mikrogliazellen können als Fresszellen des Zentralnervensystems angesehen werden. Ruhend haben sie einen kleinen Zellkörper und schlanke, verzweigte Fortsätze. Im aktivierten Zustand schwellen sie an und ziehen ihre Fortsätze ein (Färbung gegen das Protein Iba-1).
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Neuroanatomie»
Представляем Вашему вниманию похожие книги на «Neuroanatomie» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Neuroanatomie» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.