Markus Kipp - Neuroanatomie
Здесь есть возможность читать онлайн «Markus Kipp - Neuroanatomie» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на немецком языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.
- Название:Neuroanatomie
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Neuroanatomie: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Neuroanatomie»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Neuroanatomie erklärt anschaulich die wichtigsten und prüfungsrelevanten funktionellen und anatomischen Zusammenhänge des Zentralen Nervensystems. Das Buch eignet sich daher ideal zum Nachschlagen, Lernen und Verstehen in jeder Lernphase: für die Kurs- und Semesterbegleitung, fürs nächste Testat, zur effektiven Vorbereitung auf die Semesterklausur oder das Physikum. Hinzu kommt als neues Konzept die didaktische Verzahnung von schematischen Grafiken und detaillierten Fotografien von Gehirn- und Rückenmarkspräparaten. Damit werden Theorie und Praxis präzise und anschaulich miteinander verbunden und die Lerneffekte intensiviert. Die inhaltliche Gliederung sorgt für Struktur und prompten Zugriff auf den Lernstoff: • Zunächst werden die Grundlagen der Neuroanatomie vorgestellt. • Die einzelnen Bestandteile und Strukturen des Zentralen Nervensystems werden anschließend im Hauptteil erklärt. • Es folgt eine Betrachtung ihrer Zusammenhänge und ihres ineinandergreifenden Zusammenspiels. • Ein eigenes Kapitel ist den bildgebenden Verfahren und ihrer Anwendung in der Neuroanatomie gewidmet. Das Buch ist abwechslungsreich und didaktisch zielführend konzipiert: Ein übersichtliches, farbkodiertes Layout und durchdacht angeordnete Legenden ermöglichen einen raschen Überblick über den Lernstoff. Die komplexen Zusammenhänge der Neuroanatomie lassen sich mit diesem Konzept daher von Grund auf leicht verständlich erschließen und einprägen. Darüber hinaus enthält das Buch praktische Merkboxen und Hervorhebungen für spannende klinische, pharmakologische und wissenschaftliche Themen. Am Ende eines jeden Kapitels wird noch einmal gesondert auf die wichtigsten IMPP-relevanten Besonderheiten eingegangen. So kann das Gelernte auch immer an den MC-Fragen direkt getestet werden.
Neuroanatomie — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Neuroanatomie», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Axonaler Transport
Viele Substanzen, wie etwa Proteine, werden im neuronalen Zellkörper (Soma/Perikaryon) synthetisiert und von dort über einen speziellen Transportmechanismus zu ihrem Zielort (z. B. zur Synapse) transportiert: man spricht vom axonalen Transport. Vom Zellkörper zur Synapse ( anterograd, stromabwärts) werden unter anderem Membranmaterial und zur Sekretion bestimmte Substanzen (wie Neurotransmitter) transportiert. Dies geschieht über Granula oder Vesikel, die an das Motorprotein Kinesingeheftet sind ( Abb. 1.6).
Abb. 1.6
Axonaler Transport
Axone leiten nicht nur Aktionspotenziale weiter, sondern transportieren auch verschiedene intrazelluläre Substanzen (zum Beispiel Neurotransmitter oder Wachstumsfaktoren). Man unterscheidet einen Transport in gleicher Richtung wie das Aktionspotenzial (anterograder Transport, vermittelt durch Kinesin) und einen in gegensätzlicher Richtung (retrograder Transport, vermittelt durch Dynein). Viren können so von der Synapse in Richtung Soma wandern und ein Neuron dauerhaft infizieren (Herpes-Infektion).
Beim sogenannten retrograden Transport ist die Geschwindigkeit etwas geringer; hier werden Endprodukte des Stoffwechsels zurück zum Soma transportiert, außerdem zum Ab- und Umbau bestimmtes Membranmaterial sowie verschiedene Nervenwachstumsfaktoren, die für das Überleben der Nervenzelle notwendig sind. Der retrogradeTransport erfolgt über Vesikel, die an das Motorprotein Dyneingeheftet sind.
 Klinik
Klinik
Diese axonalen Transportvorgänge nutzen bestimmte Erreger aus, um sich im Gehirn einzunisten. Herpes-simplex- und Polioviren etwa gelangen durch den retrograden Transport ins Zentralnervensystem. Bei abgeschwächter Immunlage können schwerwiegende Entzündungen im Gehirn die Folge sein (Enzephalitis). Im Falle der Herpes-Viren können diese auch wieder entlang der Nervenbahnen in Richtung Haut wandern und dort zu einer lokalen Entzündung führen. Man bekommt schmerzhafte Herpesbläschen, z. B. im Bereich der Lippe.
 Forschung
Forschung
Sowohl der anterograde als auch der retrograde Transportmechanismus werden in den Neurowissenschaften genutzt, um Verbindungen im Gehirn zu verfolgen (sog. Tracingexperimente). Appliziert man spezifische Farbstoffe in eine bestimme Gehirnregion, werden diese in Nervenzellen aufgenommen und entweder anterograd oder retrograd transportiert. Mit Hilfe dieser eleganten Technik kann man feststellen, wohin ein Axon zieht (anterograder Transport) bzw. woher es kommt (retrograder Transport). Diese Technik hat wesentlich dazu beigetragen zu verstehen, welche Gehirnregionen untereinander in Verbindung stehen.
Dendriten von Nervenzellen
Das Wort „Dendrit“ leitet sich aus dem griechischen Wort für „Baum“ ab (Dendriten ähneln den Ästen eines Baumes, die vom Soma abgehen). Die Dendriten eines einzigen Neurons in ihrer Gesamtheit nennt man Dendritenbaum. Anhand der großen Vielfalt an Formen und Größen von Dendritenbäumen lassen sich die Neuronen in verschiedene Untergruppen einteilen: multipolare Neurone, bipolare Neurone, pseudounipolare Neurone, und unipolare Neurone ( Abb. 1.7).
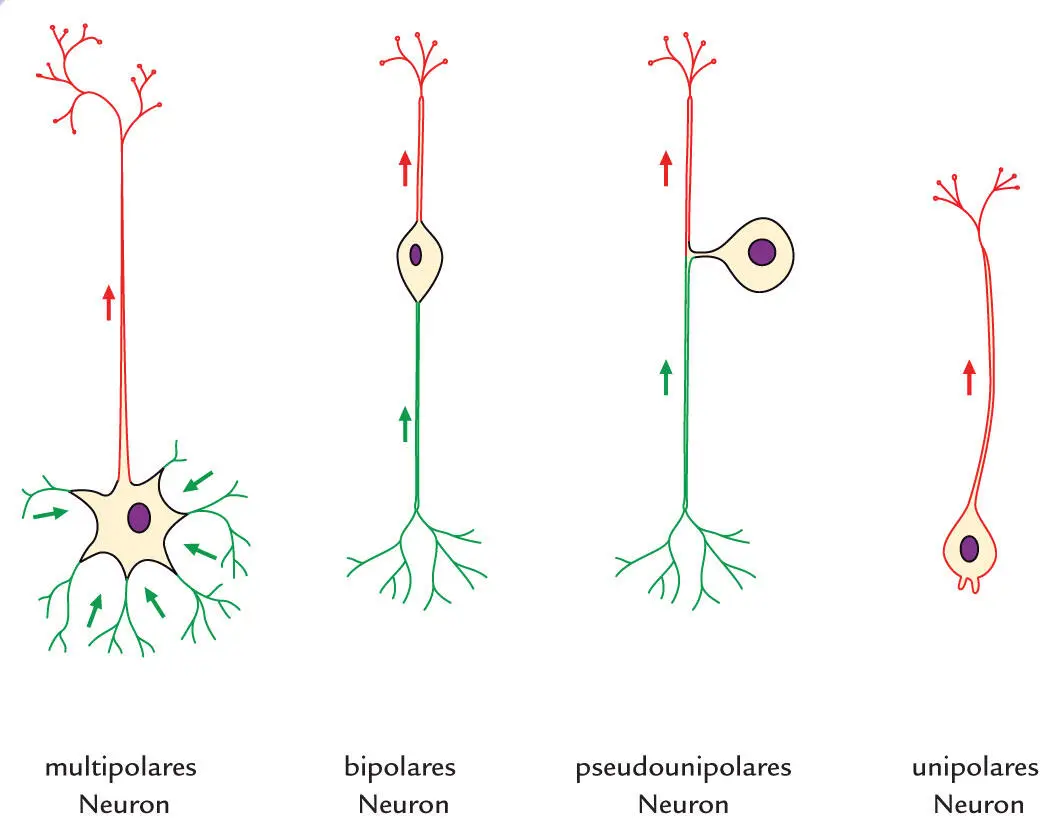
Abb. 1.7
Man klassifiziert Nervenzellen anhand der Morphologie ihrer Dendritenbäume (grün): Multipolare Neurone kommen am häufigsten vor und verfügen über ein Axon und mehr als einen Dendriten. Bipolare Neurone besitzen neben ihrem Axon genau einen Dendriten. Pseudounipolare Neurone entwickeln sich aus bipolar angelegten Neuronen. Es entspringt aus ihrem Soma zunächst nur ein Fortsatz, der sich im Verlauf in Axon und Dendrit aufzweigt. Unipolare Neurone sind sehr selten. Sie besitzen keinen Dendriten – ihre Reizwahrnehmung findet direkt am Soma oder am Axon statt.
Pseudounipolare Neurone entwickeln sich zunächst aus bipolar angelegten Zellen, deren zwei Fortsätze dann aber aufeinander zuwachsen und an den Abgangsstellen auf eine kurze Strecke miteinander verwachsen. Das Zytoplasma von Dendriten ähnelt größtenteils dem der Axone.
Die beiden Neurowissenschaftler O. Steward und W. B. Levy fanden 1982 heraus, dass in Dendriten Polyribosomen vorkommen, die häufig direkt unter den Dornfortsätzen angesiedelt sind. 2 , 3 Ihre Untersuchungen zeigten, dass die synaptische Signalübertragung in einigen Neuronen tatsächlich eine lokal begrenzte Proteinsynthese direkt im Bereich des Dendritenbaumes induzieren kann. Heute weiß man, dass diese synaptische Regulierung der lokalen Proteinbiosynthese für Lernprozesse von entscheidender Bedeutung ist.
Gliazellen
Im Rahmen der Erforschung der Ultrastruktur des Nervensystems sind den damaligen Histologen zuallererst die Nervenzellen aufgefallen. Nach und nach wurde jedoch klar, dass das Nervensystem nicht nur aus Neuronen, sondern auch aus anderen Zellen besteht, die sowohl morphologisch als auch funktional nicht so recht in das Bild der Nervenzellen passen wollten. Der Mitentdecker dieser nicht neuronalen Zellen, Rudolf Virchow, vermutete Mitte des 19. Jahrhunderts eine Stütz- und Haltefunktion und gab ihnen deshalb den Namen Gliazellen, abgeleitet aus dem griechischen Wort glia für „Leim“. Gliazelle ist also ein Sammelbegriff für strukturell und funktionell von den Neuronen abgrenzbare Zellen im Nervengewebe. Mittels unterschiedlicher Färbemethoden durch Santiago Ramón y Cajal, Pío del Río Hortega und Camillo Golgi konnten sie Ende des 19. Jahrhunderts weiter subklassifiziert werden.
Ersten Untersuchungen zufolge bilden die Gliazellen ein Stützgerüst für die Nervenzellen und sorgen für deren gegenseitige elektrische Isolation. Neuere Erkenntnisse zeigten, dass Gliazellen maßgeblich am Stoff- und Flüssigkeitstransport sowie an der Aufrechterhaltung der Homöostase im Gehirn beteiligt sind und im Prozess der Informationsverarbeitung, -speicherung und -weiterleitung mitwirken. Gliazellen sind somit unabdingbare Hilfszellen der Nervenzellen, deren spezifische Funktionen von ihnen abhängig sind. Sie besitzen Rezeptoren für viele Neurotransmitter und andere effektorische Moleküle. Es gibt sie sowohl im zentralen als auch im peripheren Nervensystem.
Eine erste Unterscheidung der Gliazellen wurde anhand ihrer Größe vorgenommen. Dementsprechend kann Mikrogliavon Makrogliaunterschieden werden. Zu den zentralen Makrogliazellen zählt man die Astrozyten, Oligodendrozyten und die Ependymzellen ( Abb. 1.8).
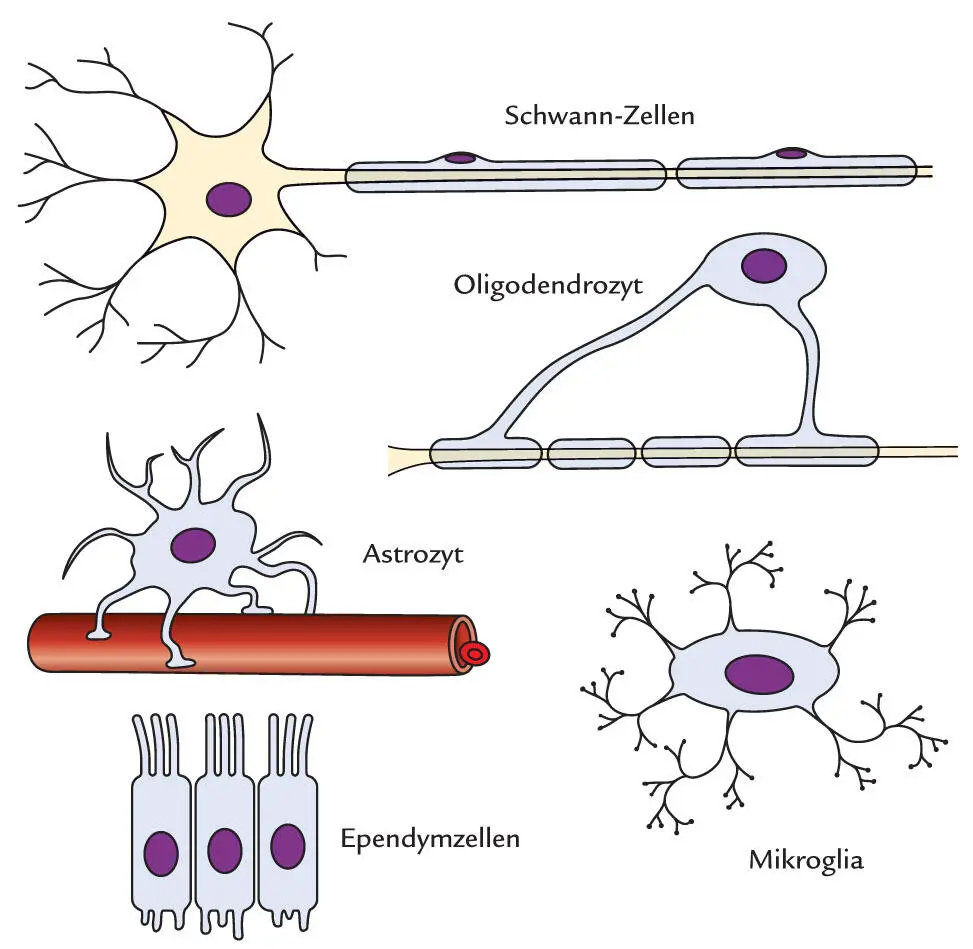
Abb. 1.8
Übersicht über verschiedene Gliazellen. Gliazellen lassen sich u. a. anhand ihrer Größe unterscheiden.
Zu den Makrogliazellen zählt man:
•Schwann-Zellen und Oligdendrozyten, die Myelinscheiden synthetisieren
•Astrozyten, die das chemische Milieu des Extrazellulärraums regulieren
•Ependymzellen, die die inneren Liquorräume auskleiden
Mikrogliazellen sind eine wichtige Effektorzellpopulation des angeborenen Immunsystems.
Bei der Unterscheidung von Mikro-und Makroglia hatten die alten Neurohistologen, ohne davon zu wissen, ein glückliches Händchen. Wie heute bekannt ist, haben Mikrogliazellen und Makrogliazellen entwicklungsgeschichtlich nichts miteinander zu tun. Makrogliazellen, also Oligodendrozyten, Astrozyten und Ependymzellen entstammen allesamt dem Neuroektoderm. Bei Mikrogliazellen handelt es sich im Gegensatz dazu um eingewanderte Blutzellen; sie entstammen also dem Mesoderm.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Neuroanatomie»
Представляем Вашему вниманию похожие книги на «Neuroanatomie» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Neuroanatomie» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.