Fabián Jaksic Andrade - Invasiones biológicas en Chile
Здесь есть возможность читать онлайн «Fabián Jaksic Andrade - Invasiones biológicas en Chile» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на испанском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Invasiones biológicas en Chile
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Invasiones biológicas en Chile: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Invasiones biológicas en Chile»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Invasiones biológicas en Chile — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Invasiones biológicas en Chile», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
La Hipótesis del Arma Novedosa fue originalmente desarrollada para explicar la naturalización de plantas que exudan compuestos alelopáticos (Thompson 1991; Callaway & Ridenour 2004). Específicamente, esta hipótesis propone que la liberación de dichos compuestos aleloquímicos por parte de la planta exótica puede actuar como factor de inhibición o exclusión del crecimiento de plantas nativas de la comunidad residente. Este hecho favorecería la naturalización al reducir a los competidores nativos, y probablemente también podrían desalentar a los herbívoros del suelo. Desde hace tiempo, la alelopatía ha sido sugerida como uno de los factores que afectan la dinámica vegetacional en ecosistemas mediterráneos y la capacidad de invadirlos (Müller 1966; Ballester & Vieitez 1979). Sin embargo, este efecto ha sido escasamente evaluado en condiciones de campo (Montenegro et al. 1978; Fuentes et al. 1984, 1987; Kruse et al. 2010). Existen pocos ejemplos que demuestran la importancia de la alelopatía para el éxito en la invasión de ecosistemas mediterráneos, como el caso de los pastos Elytrigia repens y Vulpia myuros , ambas especies de origen europeo (Friebe et al. 1995; An et al. 1997), o Centaurea diffusa , un invasor de Eurasia cuyos efectos alelopáticos se expresan sobre hierbas del oeste de América del Norte (Hierro & Callaway 2003). Particularmente severo parece ser el efecto de la lantana ( Lantana camara ), que desde América tropical y Sudamérica ha invadido bosques mediterráneos de Australia, ejerciendo efectos alelopáticos (Gentle & Duggin 1997). La evidencia experimental señala que la exudación de compuestos alelopáticos por la lantana es capaz de suprimir la germinación y reducir la tasa de crecimiento y la supervivencia de dos especies nativas (Gentle & Duggin 1997).
Finalmente, la Hipótesis de Habilidad Competitiva Incrementada se basa en que los organismos se encuentran sujetos a compromisos de asignación de los nutrientes y la energía disponible, que les permiten resolver funciones fisiológicas, tales como reproducción, crecimiento y defensa (Tilman 1988; Stearns 1992). Asumiendo que en las comunidades recipientes existen pocos enemigos naturales, la hipótesis predice que una cantidad neta de nutrientes y energía será reasignada desde funciones defensivas, por ejemplo, hacia crecimiento y reproducción. Esta hipótesis concibe a las especies naturalizadas como capaces de cierto grado de exención en los compromisos fisiológicos, cuyo resultado se traduce en un mejor –y probablemente mayor– desempeño competitivo frente a las especies nativas (Daehler & Strong 1997; Willis et al. 1999).
Interacciones facilitadoras
La mayor parte de las hipótesis y explicaciones teóricas respecto de la naturalización enfatizan las interacciones antagónicas, ya sea de competencia, depredación o parasitismo. No obstante, algunos estudios han mostrado la importancia de las interacciones positivas al interior de la comunidad (i.e., mutualismo, facilitación) en promover la naturalización (Simberloff & Von Holle 1999; Richardson et al. 2000a). Estas interacciones positivas pueden establecerse entre invasores que favorecen mutuamente su naturalización (Simberloff & Von Holle 1999; Richardson et al. 2000a; Christian 2001; Grosholz 2005), o entre especies exóticas y nativas (Richardson et al. 2000a). Uno de los ejemplos más citados en Chile supone el presunto efecto de los herbívoros exóticos tanto domésticos (caballos, vacas) como silvestres (conejos, liebres), que luego de ser introducidos en la parte central del territorio –así como en otras comunidades mediterráneas– habrían facilitado la expansión de hierbas exóticas naturalizadas en las formaciones de matorral (Kruger et al. 1989; Kalin-Arroyo et al. 2000). Efectivamente, estos herbívoros pueden consumir y defecar semillas intactas, especialmente de hierbas que tienden a ser muy pequeñas y resistentes a la digestión. Sin embargo, hasta la fecha no se han realizado estudios cuantitativos de este fenómeno.
 Biodiversidad como resistencia biótica a la naturalización
Biodiversidad como resistencia biótica a la naturalización
Elton (1958) observó que las comunidades más ricas en especies usualmente contienen menos taxa exóticos que las comunidades menos diversas. Este autor propuso que la susceptibilidad a la invasión se relaciona negativamente con la riqueza comunitaria, por lo que a medida que aumenta la riqueza se reduciría el número o proporción de especies exóticas establecidas en la comunidad ( Figura 4.4 A). Posteriormente, MacArthur (1970) le dio condición de hipótesis a la observación de Elton, argumentando que el mecanismo implícito en esta relación estaría dado por competencia interespecífica. Invocando la Teoría de Nicho, MacArthur planteó que una mayor diversidad de especies en la comunidad recipiente permitiría ocupar la mayor parte de los recursos (o ejes de nicho). A medida que la comunidad se satura, los recursos disponibles para nuevos colonizadores se verían reducidos, y con ello la probabilidad de establecerse. De acuerdo a su riqueza, las comunidades serían capaces de albergar un número determinado de especies exóticas, por lo que se dice que estas exhiben invasibilidad o resistencia diferencial (Rejmánek 1989; Levine & D’Antonio 1999; Londsdale 1999).
En los últimos 20 años se han realizado numerosos estudios para determinar la relación funcional entre la diversidad de la comunidad recipiente y la invasibilidad o resistencia a la invasión. Un hecho controversial emergió al constatar que, a escala regional (una región incluye un conjunto de comunidades), la riqueza de especies residentes mostró una relación positiva –y no negativa– con el número de especies exóticas establecidas ( Figura 4.4 B; Fox & Fox 1986; Crawley et al. 1999; Lonsdale 1999; Stohlgren et al. 1999; Levine 2000). Estos hallazgos abrieron diversos focos de discusión, uno de ellos acerca del papel de la diversidad como mecanismo efectivo y eficiente de repulsión de nuevas especies, y la importancia de la competencia interespecífica como factor de resistencia comunitaria (Levine & D’Antonio 1999).
| FIGURA 4.4 |
| Relaciones hipotéticas entre la diversidad comunitaria (constituida por especies nativas) y el éxito de las invasiones biológicas evaluado como el número de especies exóticas capaces de colonizar en la comunidad. A) Relación negativa que describe que a mayor diversidad de la comunidad recipiente, menor será la probabilidad de éxito de invasión; el mecanismo de resistencia biótica sería la competencia interespecífica (Elton 1958); B) Relación positiva que asume que un incremento en la diversidad comunitaria resultaría en un incremento de la probabilidad de éxito de invasión; este patrón estaría mediado por un aumento de la heterogeneidad espacial y de la disponibilidad de recursos a medida que aumenta la diversidad (Levine 2000); C) Visión conciliadora que describe las tendencias positivas y negativas observadas entre la riqueza comunitaria nativa y el éxito de invasión; este modelo propone que la discrepancia ha emergido como consecuencia de la escala espacial en que esta relación funcional ha sido estudiada (Shea & Chesson 2002). |
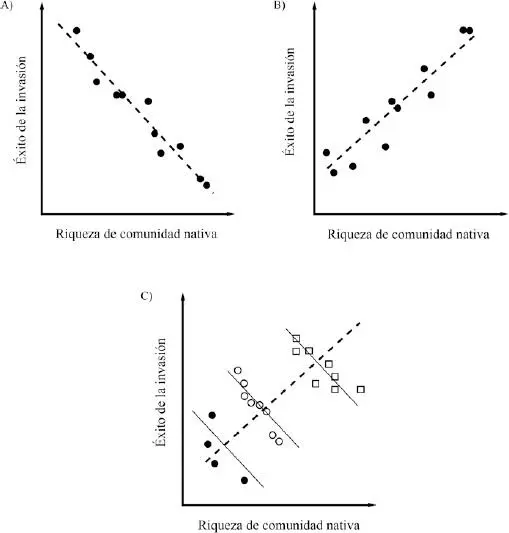 |
Desde la perspectiva regional, las comunidades se ven expuestas a un recambio natural de especies (β-diversidad) producto de la heterogeneidad de condiciones ambientales (clima, perturbaciones), geográficas y de recursos (Ricklefs 1987). Así, el conjunto de comunidades de una región conforman hábitats propicios para albergar un creciente número de especies exóticas (Naeem et al. 2000; Cornwell & Grubb 2003). Esta situación no permite reconocer si la riqueza efectivamente promueve la resistencia a la invasión y de qué manera. Para desentrañar esta interrogante se han realizado numerosos experimentos de campo y de microcosmos, los que han aportado claridad a la discusión. En términos sencillos, estos experimentos controlan el número y/o abundancia de las especies nativas, adicionando números variables de especies exóticas (o sus propágulos). Posteriormente, evalúan si la riqueza comunitaria inicial afectó el número, abundancia o biomasa de los elementos exóticos establecidos (Tilman 1997).
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Invasiones biológicas en Chile»
Представляем Вашему вниманию похожие книги на «Invasiones biológicas en Chile» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.











Обсуждение, отзывы о книге «Invasiones biológicas en Chile» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.