Naoki Sugimoto - Chemistry and Biology of Non-canonical Nucleic Acids
Здесь есть возможность читать онлайн «Naoki Sugimoto - Chemistry and Biology of Non-canonical Nucleic Acids» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Chemistry and Biology of Non-canonical Nucleic Acids
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Chemistry and Biology of Non-canonical Nucleic Acids: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Chemistry and Biology of Non-canonical Nucleic Acids»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Chemistry and Biology of Non-canonical Nucleic Acids — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Chemistry and Biology of Non-canonical Nucleic Acids», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
2.6.2.3 Internal Loop
An internal loop is formed when single-stranded loops on two strands are enclosed by adjacent two stems ( Figure 2.10). Internal loops can be symmetrical or asymmetrical with respect to the number of loop residues on each strand. In the symmetrical internal loops, there is high possibility of forming the unusual base pairs, which are described above, between the nucleobases present at opposite positions. Fully paired and stacked internal loops consisting of eight unusual base pairs have been structurally observed ( Figure 2.10) [22]. In contrast, asymmetric internal loops often become motifs that shape a three-dimensional RNA structures by bringing large changes in the helical structure such as bends and turns. Certain asymmetric internal loop motifs, such as kink-turn (K-turn) and reverse K-turn, have been identified and characterized as resulting in sharp turns important for tertiary structure formation ( Figure 2.10) [23]. By considering the possible internal loops diversities, relatively few experimental data are available for stabilities of internal loops.
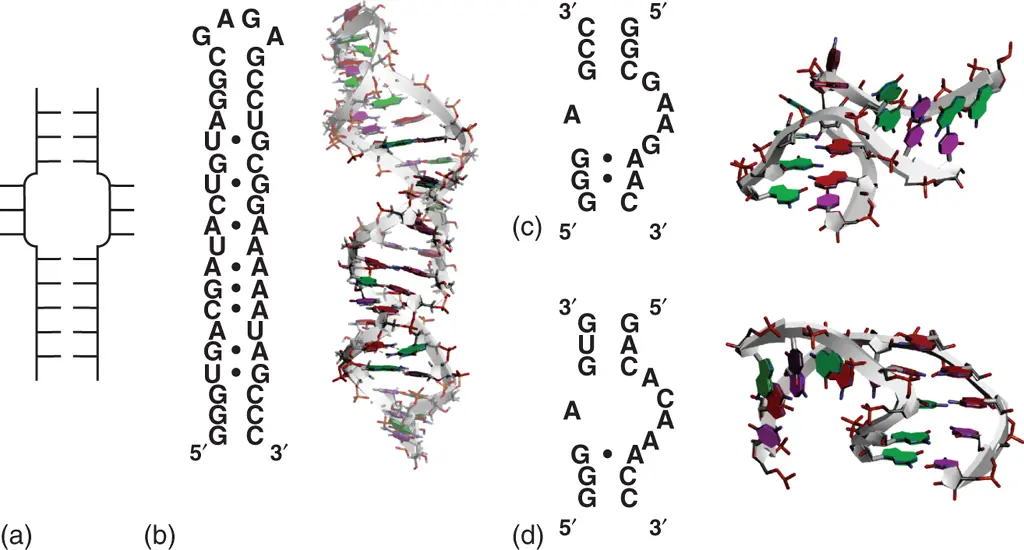
Figure 2.10 Internal loop structure. (a) General secondary structure of internal loop. (b) Sequence and tertiary structure of RNA hairpin containing internal loop forming consecutive mismatched base pairs (PDB ID: 1MNX). RNA sequence is derived from loop E region of 5S ribosomal RNA. Nucleobases forming mismatched base pairs in the internal loop, in which base pairs are shown with black circles in the secondary structure, are emphasized dark. Sequences and structures of typical kink-turn (c; PDB ID: 5FJ1) and reverse kink-turn (d; PDB ID: 1ZZN) motifs. Mismatched base pairs are shown with black circles in the secondary structures.
2.6.3 Elements in Tertiary Interactions of RNA
RNAs are commonly represented planarly as their secondary structure. However, most RNAs are folded into their defined tertiary structures that correlate with their unique functions including contribution to the gene regulation. To form a stable tertiary structure, three-dimensional interactions between secondary structure motifs including loop–loop, loop–helix, and helix–helix interactions are indispensable ( Figure 2.11) [24].
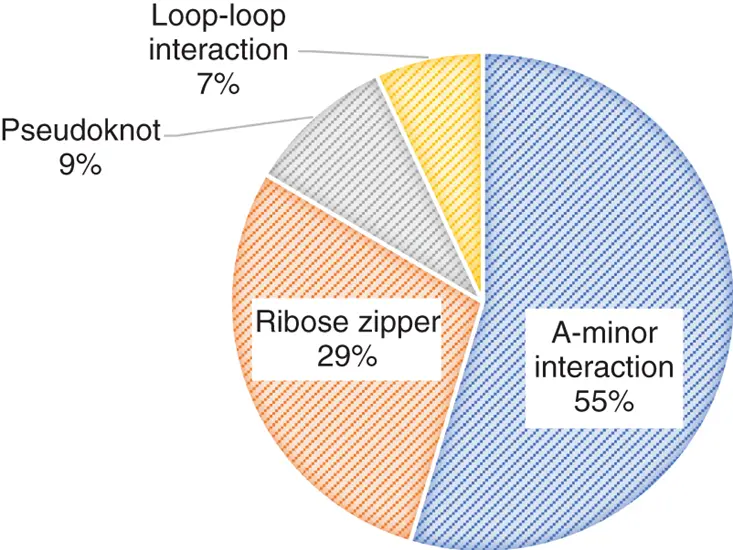
Figure 2.11 Distribution of RNA tertiary motifs except coaxial stacking of helices in the nonredundant data set of 54 high-resolution crystal structures [24a].
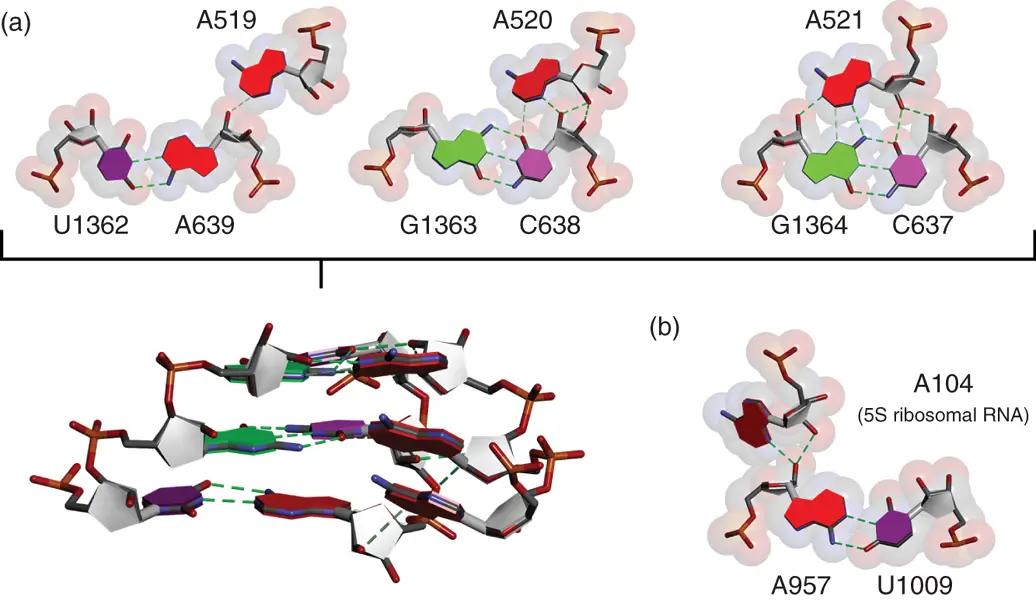
Figure 2.12 A-minor interactions. Examples of hydrogen bonding patterns of consecutive A-minor interactions observed in 23S ribosomal RNA of H. marismortui 50S ribosome (a) and that between 23S and 5S ribosomal RNAs (b) (PDBID: 1FFK). Alphabets and numbers show nucleotide positions in the ribosomal RNAs. Hydrogen bonds are shown in dashed lines. Side view of three consecutive A-minor interactions is shown below.
2.6.3.1 A-Minor Interactions
A-minor interactions are the most widely used in tertiary interaction of RNAs [24a]. These are characterized by insertion of minor groove edges of adenine nucleobases into minor groove of neighboring helices, preferentially where G·C base pairs are formed. Inserted adenine nucleobase forms hydrogen bonds with one or both of the 2′-hydroxyl group of the ribose in the base pair. There are various hydrogen bonding patterns in A-minor interactions ( Figure 2.12). Formation of hydrogen bonds between bulged adenine and minor groove of the neighboring helix contributes to the stabilization of loop–helix and helix–helix interactions [25]. Although some of the A-minor interactions, which cause tight packing of the adenine nucleobase into the minor groove, could largely contribute to the thermodynamic stability of RNA tertiary structure [26], energetic contribution of each A-minor interaction is considered to be weaker than other tertiary interaction motifs with rigid structure characteristics. It is likely that the A-minor interaction plays roles assisting in the shaping of the tertiary structure of RNAs rather than stabilizing the structure.
2.6.3.2 Ribose Zipper
Ribose zipper is also widely used interaction in RNA tertiary structures [24a]. It is usually characterized by two consecutive residues at one segment interact with two consecutive residues from another segment distant in the sequence through hydrogen bonding involving their 2′-hydroxy groups. Various classes of the ribose zipper interactions were proposed depending on the hydrogen bonding patterns [27]. In almost of the ribose zipper classes, two interacting RNA strands are in antiparallel orientation. The ribose zipper motif is often observed with other tertiary interactions such as A-minor interaction and GNRA tetraloop receptor interaction, which is described later ( Section 2.6.3.5). For example, crystal structure of group I intron [28], one of the tertiary structures from which ribose zipper interactions were proposed, contains bulge–helix and loop–helix interactions involving the ribose zipper interaction ( Figure 2.13). Both of which are associated with A-minor interaction or GAAA tetraloop receptor interaction. Since the contribution of the ribose zipper interaction on the stability of RNA tertiary structure is weaker than other tertiary interaction motifs [26, 29], as well as the A-minor interaction, ribose zipper interactions are considered to have roles supporting tertiary interactions. However, it is also considered that the ribose zipper interaction, which is provided by the 2′-hydroxy group unique for RNA ribose, is one of the indispensable factors to form complexed structures of RNAs.
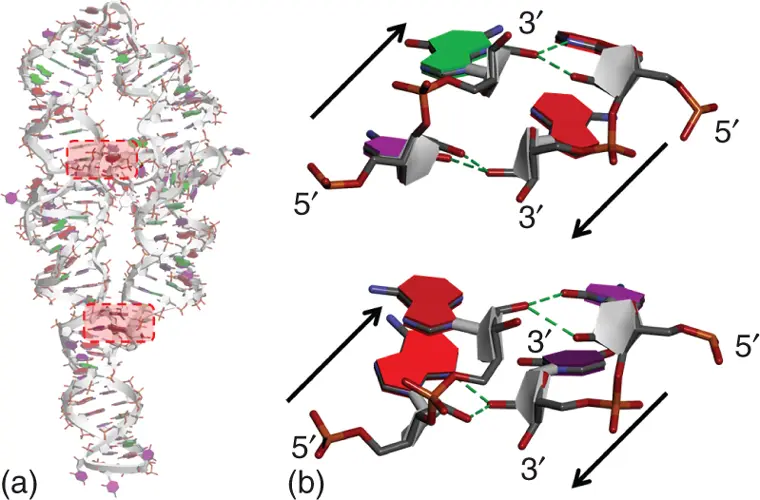
Figure 2.13 Ribose zipper interactions in tertiary structure of group I intron derived from Tetrahymena thermophila (PDBID: 1GID). Overall tertiary structure (a) and enlarged images of different types of two ribose zipper interactions (b) are shown. Hydrogen bonds involved in the ribose zipper interactions are shown in dashed lines. Arrows show orientation of RNA strands.
2.6.3.3 T-Loop Motif
T-loop motif is well-observed motif in the loop–loop interaction regions, which have been identified in a variety of RNAs [30]. T-loop motif generally consists of five consecutive nucleotides assuming a compact U-turn-like loop structure, in which the first and the fifth nucleobases form a base pair irrespective of whether it is canonical Watson–Crick or unusual one. When the T-loop motif is involved in the loop–loop interaction, the fourth and fifth nucleobases in the T-loop sandwich an extra nucleobase, which is derived from separated loop region, to make continuous base stacks. The sandwiched nucleobase usually interacts with the second nucleobase of the T-loop through hydrogen bonding [30]. In crystal structure of phenylalanyl-tRNA derived from Saccharomyces cerevisiae , in which T-loop motif was originally discovered, UUCGA loop of pentanucleotides accommodates guanine nucleobase from different tRNA loop region to form stable tertiary interaction ( Figure 2.14).
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Chemistry and Biology of Non-canonical Nucleic Acids»
Представляем Вашему вниманию похожие книги на «Chemistry and Biology of Non-canonical Nucleic Acids» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Chemistry and Biology of Non-canonical Nucleic Acids» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.