Werner Suter - Ökologie der Wirbeltiere
Здесь есть возможность читать онлайн «Werner Suter - Ökologie der Wirbeltiere» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на немецком языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Ökologie der Wirbeltiere
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Ökologie der Wirbeltiere: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Ökologie der Wirbeltiere»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Ökologie der Wirbeltiere — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Ökologie der Wirbeltiere», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
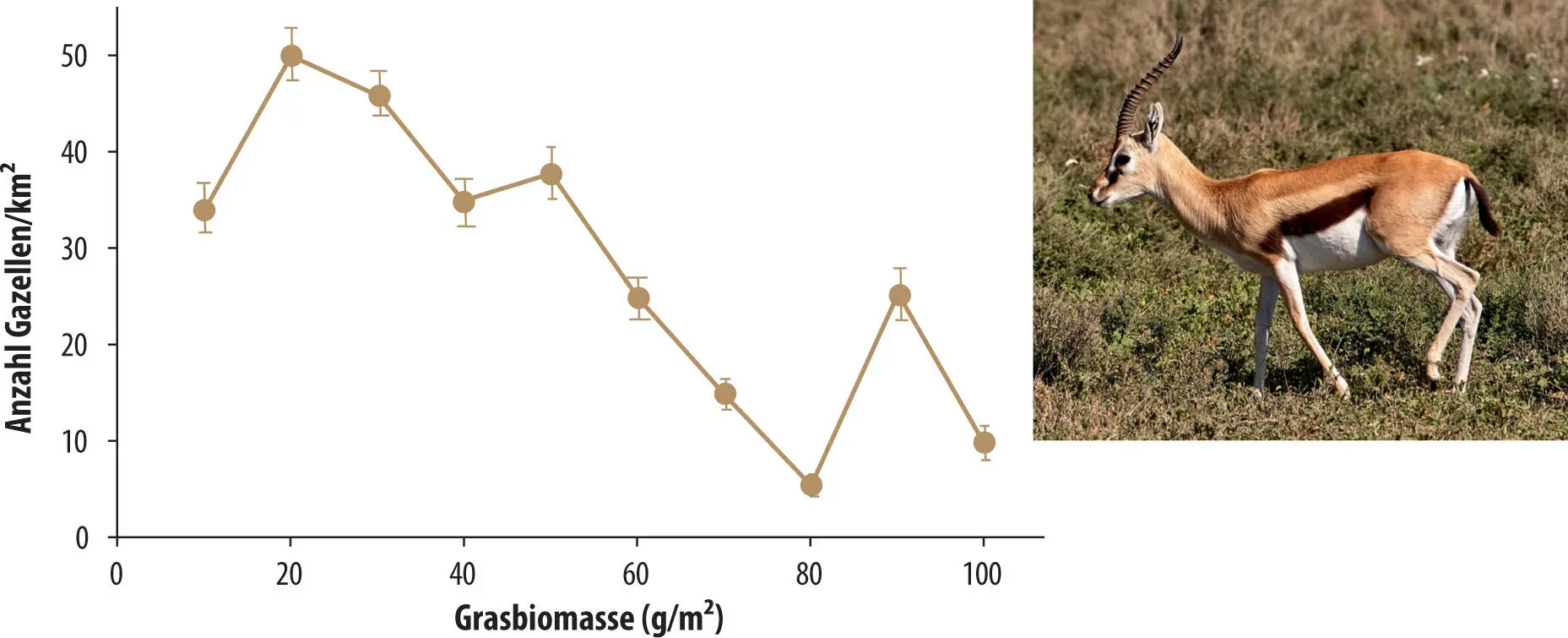
Abb. 3.15 Populationsdichten (Mittelwerte und Standardfehler) der wandernden Thomsongazelle im Serengeti-Nationalpark, Tansania (dazu Kap.6.3), sind dort am höchsten, wo die Grasbiomasse und damit die Grashöhe niedrig ist. Modellrechnungen ergaben, dass die Gazellen in niedrigem Gras die höchste Rate der Energiegewinnung erzielten (Abbildung neu gezeichnet nach Fryxell et al. 2004).
Größere Herbivoren haben die Möglichkeit, Bissgröße und -rate zu variieren; sie steigern diese bei größerem Hunger oder wenn die Zeit zur Nahrungsaufnahme limitiert ist. Bei höheren Nährstoffbedürfnissen, etwa im Falle von laktierenden Weibchen, können sie auch die gesamte Fresszeit steigern (Ruckstuhl 1998). Weshalb fressen sie dann nicht generell schneller oder in längeren Perioden? Der Grund liegt wohl bei den versteckten Kosten, etwa verminderter Feinderkennung durch geringere Aufmerksamkeit oder bei verdauungsphysiologischen Konsequenzen des Wiederkäuens ( Kap. 2.4und 2.5); auch die zum Abreißen der Pflanzen benötigten Scherkräfte müssen berücksichtigt werden (Fortin et al. 2004; Newman J. 2007; Shipley 2007).
Beweidung von patches
Auch wenn für die meisten größeren Herbivoren die Nahrung relativ flächig über die Landschaft verteilt ist, so präsentiert sich auf kleiner Skala doch meist eine deutliche Heterogenität. Man erkennt patches, im Grasland etwa Flecken einheitlicher Abundanz, Artenzusammensetzung und Qualität des Grases. Für Laubäser kann ein einzelner Baum oder Busch einen patch darstellen (Searle & Shipley 2008). Es ist deshalb nicht verwunderlich, dass die Optimal-patch-Theorie ( Kap. 3.4) auch auf Herbivoren angewandt wurde. Für Herbivoren postuliert das marginal value theorem, dass sie einen patch so lange beweiden sollten, wie die erzielte Nettoaufnahmerate über jener der Umgebung liegt. Daraus ergeben sich für Herbivoren zwei Fragen: Welcher patch soll gewählt werden, und wann soll er wieder verlassen werden? Die dazu gewonnenen Resultate sind nicht eindeutig. Dies hat einerseits damit zu tun, dass nicht alle Herbivoren stets die Nettoaufnahmerate maximieren; je nach Art und Umständen können auch patches mit hoher Biomasse, aber geringer Qualität favorisiert werden (van der Wal et al. 2000a; Bergman et al. 2001). Generell scheinen Herbivoren die für sie günstigen Flächen zu erkennen, verhalten sich aber bei der Beweidungsdauer nicht optimal; oft werden schlechtere patches «zu lange» und gute patches weniger lang als erwartet genutzt (Fryxell 2008; Bailey & Provenza 2008). Offensichtlich ist es für Herbivoren im Grasland nicht immer einfach, einen unbekannten patch nach seiner Qualität zu beurteilen und die zu erwartende Nettoaufnahmerate eindeutig abzuschätzen. Bei hoher Vertrautheit mit einem Gebiet beweiden Herbivoren einen patch eher gemäß dem Grenzwertprinzip (Focardi et al. 1996; Fryxell et al. 2014). Auch Laubäser sind mit räumlicher Hetero-genität konfrontiert; für sie stellt sich ein Baum oder größerer Busch als patch dar. Messungen an Elchen (Alces alces) zeigten, dass die Äsungsdauer an einzelnen Bäumen mit der Baumgröße zunahm, doch war die Beziehung linear und zeigte keine Abflachung, wie sie bei Annäherung an den Grenzertrag zu erwarten gewesen wäre. Tatsächlich verließen die Elche den Baum, bevor das erreichbare Laub erschöpft war (Åström et al. 1990). Dass Herbivoren einen patch meistens vor dem Erreichen des Grenzertrags aufgeben und einen neuen aufsuchen, haben viele Untersuchungen gezeigt. Offensichtlich ist das Gewinnen von Information über alternative patches von großer Bedeutung; dies geschieht vorzugsweise durch Probeweiden und nicht allein durch eine optische Beurteilung (Illius & Gordon 1993).
Funktionelle Reaktionen
Eine funktionelle Reaktion (functional response) ist ein Modell dafür, wie die Aufnahmerate eines Konsumenten mit zunehmender Häufigkeit der Nahrung ansteigt. Von C. S. Holling (*1930), einem kanadischen Entomologen, wurden drei Typen (I, II und III) beschrieben, die sich zunächst auf Prädatoren und Parasiten bezogen (Holling 1959; Kap. 9.2). Bei Typ 1 nimmt die Aufnahmerate mit der Abundanz der Nahrung linear bis zu einem oberen Limit zu. Bei Typ II flacht die Aufnahmerate kurvilinear mit zunehmender Abundanz ab, um sich dem oberen Limit asymptotisch anzunähern. Bei Typ III ist die Funktion zusätzlich S-förmig, weil die Aufnahmerate bei geringer Nahrungsdichte ebenfalls abgeflacht ist, etwa bei versteckter Beute. Typen II und III ergeben sich, wenn die Zeit für das Handling nicht null ist. Diese Modelle können auch für Herbivoren Anwendung finden. Oftmals wurde bei Grasfressern eine Beziehung zwischen der Abundanz der Vegetation und der Aufnahmerate beobachtet, die Typ II entspricht. Bei Laubäsern ergab sich ein entsprechender Zusammenhang erst, wenn die Aufnahmerate nicht mit der Abundanz der Nahrung, sondern der realisierten Bissgröße in Beziehung gesetzt wurde. Deshalb passten Spalinger & Hobbs (1992) das Modell für die Situation der Herbivoren an, da bei diesen im Gegensatz zu Prädatoren das Handling eines Bisses, das heißt das Kauen, und die Suche für den nachfolgenden Biss zeitlich überlappen können. Eine Funktion, bei der mit großer Dichte des Angebots die Aufnahmerate wieder zurückgeht, wurde später als Typ IV beschrieben und auch bei kleineren Herbivoren gefunden (Heuermann et al. 2011).
Die Spalinger-Hobbs functional response hat sich als fruchtbare Hilfe erwiesen, die Nahrungsaufnahme durch Herbivoren auf kleinen Skalen – räumlich wie zeitlich – zu modellieren. Bissgröße und -rate konnten mit der Abundanz und räumlichen Verteilung sowie strukturellen und qualitativen Eigenschaften der Pflanzen in Verbindung gesetzt werden. Auch die Wirkung von constraints, etwa der weiter oben genannten Notwendigkeit der Wachsamkeit gegenüber Feinden, welche die Aufnahmerate verringern kann, ließen sich berücksichtigen. Es zeigte sich, dass Herbivoren ungeachtet ihrer Körpergröße in ähnlicher Weise auf die Heterogenität der Nahrungsressourcen reagieren (Hobbs et al. 2003). Weiteres zum Konzept und zur Modellierung von funktionellen Reaktionen der Herbivoren findet man in den Übersichten von Owen-Smith (2002), Illius (2006), Laca (2008), Fryxell (2008) und Searle & Shipley (2008).
Weiterführende Literatur
Auf weiterführende Literatur zu einzelnen Themen, vor allem stark auf Theorie und Modelle ausgerichtete Beiträge, wurde am Ende von verschiedenen Unterkapiteln bereits hingewiesen. Hier folgt nochmals eine Übersicht über (vorwiegend) neuere Bücher, die der Verhaltensökologie des Nahrungserwerbs ausführlich Raum widmen. Dies ist zunächst in einigen verhaltensbiologischen Standardwerken der Fall, auch wenn Aspekte der Nahrungssuche dort über verschiedene Kapitel verteilt sind:
• Alcock, J. 2013. Animal Behavior. An Evolutionary Approach. 10th ed. Sinauer Associates, Sunderland.
• Davies, N.B., J.R. Krebs & S.A. West. 2012. An Introduction to Behavioural Ecology. 4th ed. Wiley-Blackwell, Chichester.
• Kappeler, P.M. 2012. Verhaltensbiologie. 3. Aufl. Springer-Verlag, Berlin.
Zwei neue editierte Werke mit konsequent evolutionsbiologischer Ausrichtung widmen dem Nahrungserwerb hingegen eigene Kapitel:
• Danchin, E., L.-A. Giraldeau & F. Cézilly. 2008. Behavioural Ecology. Oxford University Press, Oxford.
• Westneat, D.F. & C.W. Fox (eds.). 2010. Evolutionary Behavioral Ecology. Oxford University Press, Oxford.
Ganz auf die Nahrungssuche fokussieren mindestens drei Werke, zwei von ihnen mit stark theoretischer Ausrichtung, das jüngste eine sehr ergiebige Kombination von Theorie und empirischen Befunden:
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Ökologie der Wirbeltiere»
Представляем Вашему вниманию похожие книги на «Ökologie der Wirbeltiere» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Ökologie der Wirbeltiere» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.