Werner Suter - Ökologie der Wirbeltiere
Здесь есть возможность читать онлайн «Werner Suter - Ökologie der Wirbeltiere» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на немецком языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Ökologie der Wirbeltiere
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Ökologie der Wirbeltiere: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Ökologie der Wirbeltiere»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Ökologie der Wirbeltiere — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Ökologie der Wirbeltiere», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Anpassungen einer Suchstrategie an die Prädationsgefahr können auf verschiedene Weise erfolgen, zum Beispiel durch zeitliche Verschiebungen der Nahrungssuche, durch (oftmals subtile) Änderungen in der Habitatnutzung, mittels Reduktion der Fortbewegung oder einfach durch erhöhte Wachsamkeit (Bednekoff 2007; Ferrari et al. 2009). Pinguine etwa vermeiden es, bei geringem Licht Nahrung zu suchen, obwohl sie das könnten (Ainley & Ballard 2012). Kleinsäuger nutzen hingegen geringe Lichtstärken aus: Rennmäuse (Gerbillus sp.) passen ihre Aktivitätsphasen nicht nur den Habi-tatstrukturen und der Nahrungsdichte, sondern auch dem Mondlicht an. Sie sind bei Neumond aktiver, und die Intensität ihrer Wachsamkeit ändert im Verlauf einer Nacht in Abhängigkeit von Licht und der Aktivität der Prädatoren (Kotler et al. 2002; Berger-Tal et al. 2010; Embar et al. 2011). Eine Metaanalyse von Arbeiten, die mehrheitlich an kleinen Nagetieren durchgeführt worden waren, zeigte die generelle Bedeutung von Habitatstrukturen auf: Die giving-up densities waren höher auf patches mit schlechteren Deckungsmöglichkeiten (Verdolin 2006). Dieselbe Beziehung wurde auch bei Primaten gefunden (Abb. 3.8).
Unterschiedliche Prädationsgefahr in Abhängigkeit der Habitatstrukturen, und damit räumlich unterschiedliche Kosten bei der Nahrungssuche, kann die Raumnutzung von Tieren stark beeinflussen. So gesehen, bewegen sie sich in einer landscape of fear (Laundré et al. 2001), in der die «Täler» und «Berge» Flächen mit niedrigen und hohen prädationsbedingten Kosten repräsentieren (Abb. 3.9). Genauer betrachtet, sind diese Landschaften eher landscapes of risk (Norum et al. 2015). Die Verteilung von Flächen hohen Risikos und Refugien ist abhängig von der Heterogenität der Landschaft; in gleichförmig strukturierten Landschaften kann ein Mangel an Refugien dazu führen, dass die nahrungsreichsten Flächen trotz hoher Prädationsgefahr genutzt werden (Schmidt & Kuijper 2015). Die tatsächliche Nutzung der Landschaft durch Herbivoren braucht in artenreichen Prädatoren-Beute-Gemeinschaften die Prädationsgefahr nicht genau widerzuspiegeln (Thaker et al. 2011). Verhaltensänderungen (räumliche Verlagerungen, verkürzte Verweildauern auf patches, verstärktes Wachsamkeitsverhalten auf Kosten der Nahrungsaufnahme) der Beutearten differieren, je nachdem, ob mit Ansitzjägern oder Verfolgungsjägern zu rechnen ist (Preisser et al. 2007; Wikenros et al. 2015). Im Zuge der Wiedereinbürgerung von Wölfen in nordamerikanischen Nationalparks passten die Wapitis (Cervus canadensis) ihre Raumnutzung in beschränktem Maß an diesen Verfolgungsjäger an; sie nutzten die offenen, grasbestandenen Flusstäler weniger und hielten sich vermehrt in den weniger nahrungsreichen Waldbeständen auf (Creel et al. 2005; Fortin et al. 2005; Kauffman et al. 2010). Welches allerdings die Kosten von solchen non-consumptive effects sind, ist umstritten. Befunde, dass durch den Wolf ausgelöste Risikoeffekte bei Wapitis die Energieaufnahme der Hirsche und letztlich die Geburtenrate sinken ließen (Creel et al. 2007; Christianson & Creel 2010), konnten im Rahmen umfassenderer Studien nicht bestätigt werden. Absolut gesehen, war die Chance des Zusammentreffens von Wapitis und Wölfen selbst in relativ dicht von Wölfen besiedelten Gebieten gering, denn im Mittel kam ein Wapiti lediglich alle 9–50 Tage einem Wolf auf weniger als 1 km nahe (Middleton et al. 2013).
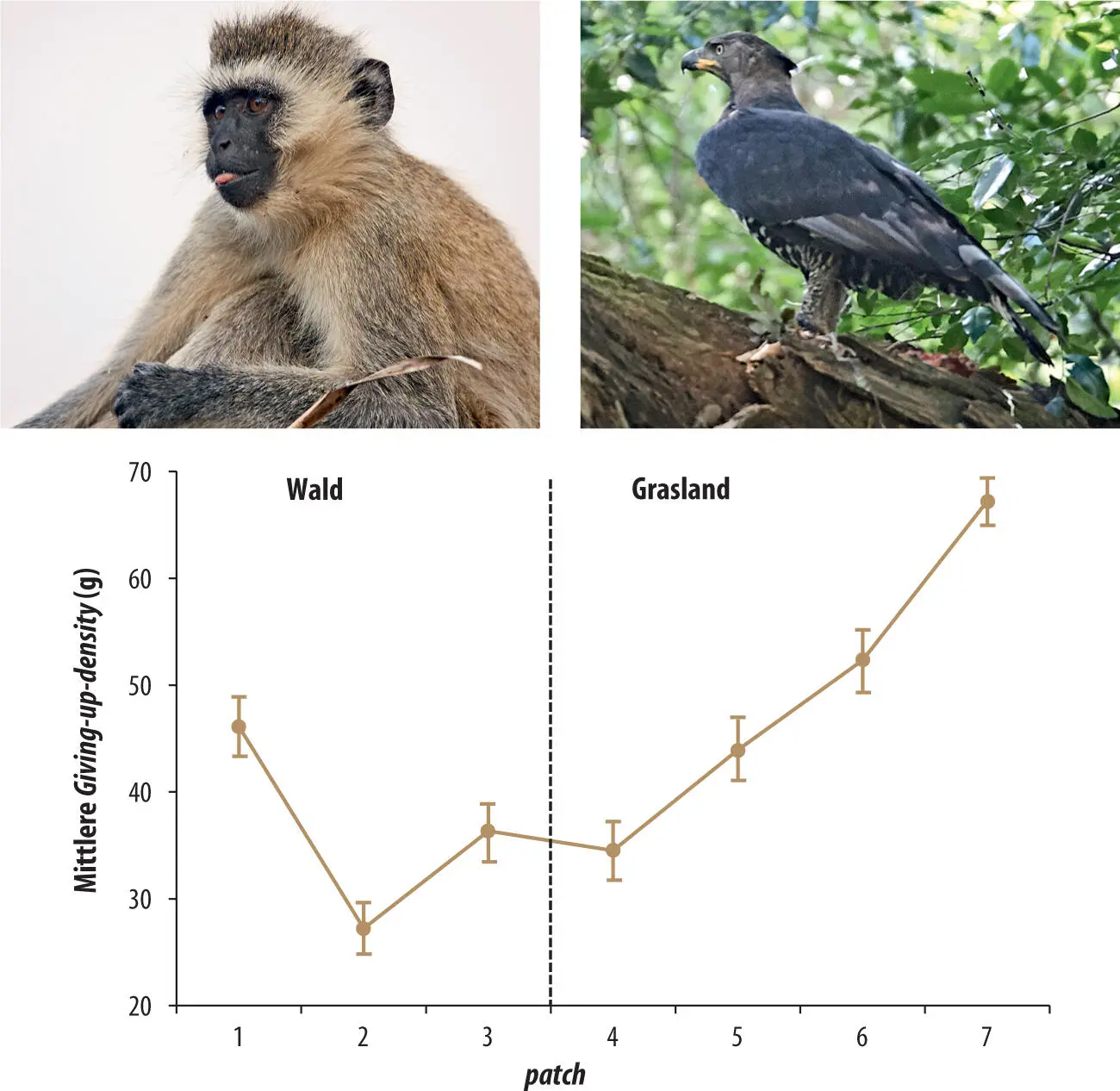
Abb. 3.8 Auch für Grünmeerkatzen (Chlorocebus aethiops) führt die Prädationsgefahr zu Kosten, welche die Maximierung der Energieaufnahme verhindern. Wild lebenden Meerkatzen wurden patches von Nahrung in Form von Futterkübeln mit Erdnüssen offeriert, die nur mit gewissem Aufwand herausgeklaubt werden konnten. Die Kübel waren auf Transekten angeordnet, die vom Baumwipfel hinunter zum Stammfuß und dann aus dem Wald hinaus in offenes Grasland führten. Prädationsgefahr ging in Wipfelnähe (patch 1) von Kronenadlern (Stephanoaetus coronatus) und am Boden (patches 4–7) von terrestrischen Prädatoren aus. Tatsächlich war die giving-up-density in mittlerer Baumhöhe (patch 2) am niedrigsten und nahm im Grasland mit zunehmender Entfernung vom Waldrand schnell zu, was bedeutet, dass die Kübel umso weniger geleert wurden, je größer die Gefahr durch Feinde war (Makin et al. 2012). Große Adler sind übrigens in allen tropischen Gebieten die Hauptprädatoren kleinerer und mittelgroßer Primaten (siehe auch Kap. 3.6) (Abbildung neu gezeichnet nach Makin et al. 2012).
«Risikolandschaften» existieren auch für Prädatoren, wenn sie selbst als Beute größerer Prädatoren infrage kommen ( Kap. 9.11). Übrigens formt auch die Präsenz des Menschen bei vielen Tieren eine landscape of fear, besonders wo der Mensch über die Jagd ebenfalls als direkter Prädator auftritt. Die anthropogene Beeinflussung des raum-zeitlichen Verhaltens kann dabei jene der natürlichen Prädatoren deutlich übertreffen (Ciuti et al. 2012; Crosmary 2012) und auch bei großen Prädatoren die Raumnutzung entscheidend gestalten (Oriol-Cotterill et al. 2015).
3.6 Nahrungssuche in der Gruppe
Bei der Großzahl der Carnivoren und vielen anderen Arten sind die Individuen einzeln unterwegs, wenn sie Nahrung suchen. Dies war die Prämisse in den bisherigen Kapiteln zur Nahrungssuche: Die Optimierungsmodelle behandelten die Tiere als ökonomisch unabhängige Einheiten. Zahlreiche Arten aber handeln anders: Die Individuen leben sozial und suchen gemeinsam Nahrung, sei es in kleinen Gruppen, sei es in großen Schwärmen oder Herden (social foraging). Betrachten wir ein Beispiel, etwa einen Schwarm von Strandläufern, kleinen Watvögeln (Limikolen), die irgendwo am Meeresstrand Nahrung suchen. Sie bewegen sich schnell vorwärts, picken hier und dort, und immer wieder kommt es während Sekundenbruchteilen zu aggressiven Gesten zwischen Individuen. An einem patch mit erhöhter Nahrungsdichte hält der Schwarm an, und einige Strandläufer erbeuten einen Ringelwurm, wobei die erfolglosen Nachbarn versuchen, ein Stück davon zu stehlen. Immer auch mustern einige der Vögel die Umgebung, und als ein Greifvogel erscheint, fliegt der ganze Schwarm unvermittelt auf. Nun, auch sozial lebende Tiere suchen ihre Nahrungsaufnahme zu optimieren. Wäre es für den einzelnen Strandläufer deshalb nicht vorteilhafter gewesen, er hätte den reichen patch allein ausbeuten können? Hätte er ihn allein gar nicht gefunden? Oder ist er durch den erfolgreichen Versuch, ein Stück Ringelwurm zu stehlen, allenfalls mit geringerem Aufwand zu Beute gekommen? Hätte er auf sich gestellt den herannahenden Prädator gesehen?

Abb. 3.9 Eine landscape of fear des Kapborstenhörnchens (Xerus inauris) in Südafrika, bei der die Kosten des Prädationsrisikos mittels Isolinien von Opportunitätskosten kartiert sind. Die Isolinien bezeichnen die Energieaufnahmerate in Joule pro Minute, bei der die Erdhörnchen ihre Nahrungssuche auf einem patch aufgaben. Je weiter weg vom Bau ein patch war, desto höher war die bei Aufgabe verbliebene Nahrungsmenge, doch formten auch Bäume und andere das Prädationsrisiko beeinflussende Elemente die Karte mit (nach van der Merwe & Brown 2008) (Abdruck mit freundlicher Genehmigung von Oxford University Press, © OUP).
Diese Fragen zu beantworten und dahinter die Gründe für die Vorteile der Nahrungssuche im sozialen Verband zu suchen, ist mit den bisherigen Optimierungsmodellen nicht möglich. Jedes Individuum hat ja das Verhalten seiner Gruppenmitglieder zu berücksichtigen und ist in seiner Entscheidungsfindung nicht mehr frei, sondern auf den Konsens in der Gruppe angewiesen (Conradt & Roper 2005). Es ist die Spieltheorie, die uns die Werkzeuge in die Hand gibt, mit denen wir Taktiken von Gruppenmitgliedern modellieren und Entscheidungen voraussagen können, um sie am realen Verhalten der Tiere zu testen. Die Modelle kommen zu dem Schluss, dass die beste Taktik eines Individuums immer von den Taktiken abhängen wird, die andere Gruppenmitglieder anwenden. Gibt es aber Strategien, die konstant besser sind als andere? Diese Strategien müssten durch die natürliche Selektion favorisiert sein und, falls sie von der ganzen Population angewendet werden, nicht durch eine Alternative verdrängt werden können – sie sind evolutionär stabil (evolutionary stable strategy, ESS; Maynard Smith 1982). Eine ESS braucht kurzfristig nicht unbedingt die vorteilhafteste Strategie für die Population zu sein; die Stabilität ist das entscheidende selektionierte Merkmal. Übersichten zur Theorie des social foraging offerieren Giraldeau & Caraco (2000), Waite & Field (2007), Giraldeau (2008b) oder Hamilton I. M. (2010). An dieser Stelle wollen wir uns nicht mit den Modellen, sondern empirischen Befunden zu den Vorteilen und Nachteilen der Nahrungssuche in der Gruppe beschäftigen; auch dazu gibt es eine vertiefte Darstellung (Krause & Ruxton 2002).
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Ökologie der Wirbeltiere»
Представляем Вашему вниманию похожие книги на «Ökologie der Wirbeltiere» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Ökologie der Wirbeltiere» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.