Armand Leroi - Mutants
Здесь есть возможность читать онлайн «Armand Leroi - Mutants» весь текст электронной книги совершенно бесплатно (целиком полную версию без сокращений). В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: New York, Год выпуска: 2005, ISBN: 2005, Издательство: Penguin Books, Жанр: История, Биология, sci_popular, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.
- Название:Mutants
- Автор:
- Издательство:Penguin Books
- Жанр:
- Год:2005
- Город:New York
- ISBN:0-670-03110-0
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Mutants: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Mutants»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
gives a brilliant narrative account of our genetic code and the captivating people whose bodies have revealed it—a French convent girl who found herself changing sex at puberty; children who, echoing Homer’s Cyclops, are born with a single eye in the middle of their foreheads; a village of long-lived Croatian dwarves; one family, whose bodies were entirely covered with hair, was kept at the Burmese royal court for four generations and gave Darwin one of his keenest insights into heredity. This elegant, humane, and engaging book “captures what we know of the development of what makes us human” (
).
Mutants — читать онлайн бесплатно полную книгу (весь текст) целиком
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Mutants», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
And many people are born with extra digits. About i in 3000 Europeans is born with extra fingers or toes (or both), and about 1 in 300 Africans. Any digit can be duplicated, but in Africans it usually a little finger (pinkie), while in Europeans it tends to be a thumb. Polydactyly is usually genetic, frequently dominant, and can run for many generations in families. Long before Gregor Mendel ever lived, the French mathematician Pierre-Louis Moreau de Maupertuis (1698–1758) described the inheritance of Polydactyly in the ancestors and descendants of a Berlin physician called Jacob Ruhe. Ruhe’s grandmother had six fingers on each hand and six toes on each foot, as did his mother, as did he and three of his seven siblings, and two of his five children. Others have claimed even more impressive polydactylous pedigrees. In 1931 the Russian geneticist E.O. Manoiloff published an account of a polydactylous Georgian, Via?eslav Michailovi? de Camio Scipion, who, he said, was able to document his descent from a lineage of polydactylous forebears reaching back six centuries.
If the apical ectodermal ridge ensures that our limbs grow out into space, another equally unobtrusive piece of limb-bud ensures that we have the right number and kinds of fingers. It was again John Saunders, along with a collaborator, Mary Gasseling, who discovered it. They found that if they transplanted a piece of mesoderm from the tailmost edge of one chicken limb-bud onto the headmost edge of another (so that the bud had two tailmost edges in opposite orientation to each other), the result was a chicken wing with twice the usual number of digits. Most remarkably of all, the experimental wings were like a particularly exotic variety of polydactyly in humans. They resembled people who, far from having just an extra digit or two, have hands and feet that are almost completely duplicated with up to ten digits each. The polydactylous wings had a peculiar mirror-image geometry, one shared by duplicated hands in humans. If each finger is given a code in which the thumb is 1, forefinger 2, index-finger 3, ring-finger 4, and pinkie 5, then a normal, five-fingered, hand has the formula ‘12345’, while a duplicated hand has the formula ‘5432112345’. It is that strangest of things, an anatomical palindrome.
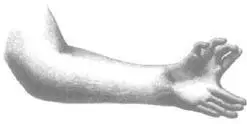
Saunders and Gasseling called their potent piece of mesoderm the ‘zone of polarising activity’ or ‘ZPA’. It is thought to be the source of a morphogen. At its source, where it is most concentrated, this morphogen induces naive mesoderm to become the little finger; further away, lower concentrations induce the ring, index, and forefinger in succession, and at the far opposite end of the limb, you get a thumb.
This account of how most of us come by our five fingers brings to mind the organiser. Like the organiser, the ZPA has the uncanny ability to impose order on its surroundings. And, just as the organiser morphogen was so eagerly sought for so long, so too, in recent years, has been the morphogen of the ZPA. It is almost certainly a signalling protein, likely a familiar one, a member of one of the great families of signalling proteins that also work elsewhere in the embryo. But limb-buds contain a plethora of such proteins, and it is hard to know which of them is the morphogen itself. In the past few years, several candidate molecules have been said to fit the bill. One of them is sonic hedgehog.
Sonic hedgehog appears in the limb-bud precisely where one would expect a morphogen to be: only in the mesoderm of the tailmost edge, exactly coincident with Saunders and Gasseling’s ZPA. It also does what one would expect a morphogen to do: shape limbs. Chicken wings can be sculpted into new and improbable forms – including duplicate mirror-image polydactylous ones – simply by manipulating the presence of sonic in the bud. And then there are the mutants. Mutations in at least ten genes cause Polydactyly in humans and all seem to affect, in some way or other, sonic’s role in the limb.
But, as we saw in the previous chapter, sonic hedgehog does not just determine how many fingers and toes we have. It also divides our brains, decides how widely spaced our eyes will be, and regulates much else besides. It is an incorrigibly promiscuous molecule. Could we see the pattern of the sonic hedgehog gene’s activity over time, as in time-lapse photography, we would see it flashing on and off throughout the developing embryo and foetus, now in this incipient organ, now in that one.
The devices responsible for all this have a formidable task, and nowhere, given sonic’s power to direct the destiny of cells, do they have much room for error. These devices are transcription factors or ‘molecular switches’. Some of them keep sonic in check. Should they be disabled by mutation, sonic turns on in parts of the limb-bud that it otherwise would not – and the result is extra fingers and toes. Other mutations do not disable the transcription factors themselves, but rather delete the regulatory elements to which they bind. The result, however, is the same: a confusion of morphogen gradients and an embarrassment of digits.
Polydactyly mutations relax control of sonic hedgehog altering the balance of power in favour of ubiquity. But other mutations have exactly the opposite effect and prevent sonic from appearing in the limb-bud at all. The most blatant example of such a mutation is, of course, one that disables the sonic gene itself. Sonic-less mice have, in addition to their many other defects, no paws. This is strikingly reminiscent of a disorder that we have already come across: acheiropody, the disorder of the aleijadinhos. Indeed, there is some (disputed) evidence that the acheiropody mutation disables a regulatory element essential to sonic’s presence in the limb.
This catalogue of mutations only hints at the complexities of gene regulation in the embryo. Whether or not a gene is turned on in a given cell depends on what transcription factors are found in that cell’s nucleus, and their presence depends on the presence of yet other transcription factors, and so on. At first glance hierarchies of this sort seem to involve us in an infinite regression in which the burden of producing order is merely placed upon a previous set of entities which must, themselves, be ordered. But this dilemma is more apparent than real. The embryo’s order is created iteratively. Sonic’s precise presence in the ZPA is defined in part by the activity of Hox genes in the trunk mesoderm from which limbs grow. But the geometrical order that these genes give to the limb is crude; sonic’s task is to refine it further. Beyond sonic there are, of course, yet further levels of refinement in which order is created on ever smaller scales, and each of them requires subtle and interminable negotiations, the nature of which we scarcely understand.
This vision of successive layers of negotiation and control may seem unimaginably complex. But in truth it is not complex enough, for it fails to capture one of the most pervasive properties of the embryo’s programme: its non-linearities. I argued that the acheiropody mutation causes a failure of sonic to appear in the limb. And yet I began this chapter by arguing that infants with amputations in the womb, of whatever severity, were due to failures of the apical ectodermal ridge and the fibroblast growth factors they produce. This may seem like a contradiction, but it is only one if we think of the various limbs’ signals as being independent of each other, when in fact they are not. For one of the most vital roles of sonic hedgehog is to maintain and shape the apical ectodermal ridge and its fibroblast growth factors; and one of the most vital roles of the apical ectodermal ridge is to maintain and shape the production of sonic hedgehog in the zone of polarising activity. There is a reciprocal flow of information as precarious as the flow of batons between two jugglers standing at opposite ends of a stage. Reciprocity of this sort is ubiquitous in the embryo and it alters the way we think about its growth and development. We begin with notions of linear pathways of command and control and simple geometries – and then watch as they unravel. For when, as in the limb, we actually begin to see the outlines of the embryo’s programme, it invariably turns out to resemble a tangle of circuits that loop vertiginously across time and space. Circuits which, in this case, ensure that when we count our fingers and toes we usually come up with twenty.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Mutants»
Представляем Вашему вниманию похожие книги на «Mutants» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Mutants» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.