An Introduction to Molecular Biotechnology
Здесь есть возможность читать онлайн «An Introduction to Molecular Biotechnology» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:An Introduction to Molecular Biotechnology
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
An Introduction to Molecular Biotechnology: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «An Introduction to Molecular Biotechnology»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
An Introduction to Molecular Biotechnology — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «An Introduction to Molecular Biotechnology», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
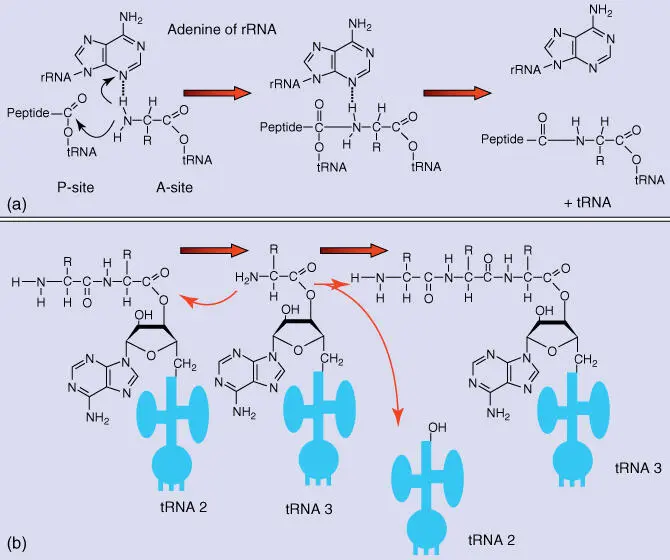
Figure 4.25 rRNA‐catalyzed peptide transfer in ribosomes. (a) Possible reaction mechanism with an adenine residue of the rRNA participating in catalysis. (b) Reaction pathway of peptidyl transfer.
After protein synthesis, the newly synthesized proteins fold themselves into the correct conformation, aided in many cases by chaperones(e.g. diverse heat shock proteins; hsp60 and hsp70 and others) acting as auxiliary enzymes. Incorrectly folded or incorrectly synthesized proteins (e.g. protein fragments resulting from strand breaking) are coupled with the protein ubiquitinand are broken down in a cellular “shredder” – the proteasomes.
Protein biosynthesis can occur on free ribosomes in the cytoplasm or on ribosomes, which bind to the rough ER (see Chapter 5). A single mRNA can be used by several ribosomes concomitantly; such structures are called polyribosome.
Prokaryotic and eukaryotic ribosomes are constructed according to a very similar pattern( Figure 4.22), and protein biosynthesis is conducted according to very similar principles. However, the particular rRNAs and ribosomal enzymes exhibit important differences. The importance of many antibiotics depends on these differences to specifically inhibit prokaryotic ribosomes. Many antibioticsintervene in bacterial protein biosynthesis (Table 4.7).
Table 4.7 Protein biosynthesis in bacterial ribosomes as a target for antibiotics.
| Antibiotic | Mode of action |
|---|---|
| Tetracycline | Inhibits A‐site in ribosomes |
| Aminoglycosides (streptomycin) | Disturbs anticodon–codon recognition and chain elongation |
| Erythromycin | Binds to 50S subunit, blocks exit site (E), and inhibits chain elongation |
| Chloramphenicol | Binds to 50S subunit and inhibits peptidyl transfer |
| Puromycin | Induces a premature chain termination |
Owing to their selectivity toward bacteria, antibiotics (which came on the market only 70 years ago) are generally substances with few side effects in humans. The search for new and more effective antibiotics is still one of the most important challenges of biotechnology and medicine because many pathogens have become resistant (overexpression of ABC transporters, target site mutations) to existing antibiotics ( multidrug‐resistant ( MDR ) pathogens). A number of pathogenic strains of Staphylococcus aureus that have become resistant to most antibiotics (so‐called methicillin‐resistant S. aureus [MRSA]) are particularly dangerous (see Section 3.2).
Further Reading
1 Alberts, B., Johnson, A., Lewis, L. et al. (2015). Molecular Biology of the Cell, 6e. New York: Garland Science.
2 Alberts, B., Bray, D., Hopkin, K. et al. (2019). Essential Cell Biology, 5e. New York: Garland Science.
3 Krebs, J., Goldstein, E.S., and Kilpatrick, S.T. (2018). Lewin's Genes XII. Burlington: Jones & Bartlett Learning.
5 Distributing Proteins in the Cell (Protein Sorting)
Michael Wink
Heidelberg University, Institute of Pharmacy and Molecular Biotechnology (IPMB), Im Neuenheimer Feld 329,, 69120 Heidelberg, Germany
The cellular compartmentswere introduced in Chapter 3. All compartments are enclosed by a biomembrane and contain a multitude of proteins. In many cases, the separation of proteins in a cell is compartment specific, meaning that every compartment harbors its own set of proteins. Every animal cell contains about 10 10single protein molecules, whose synthesis begins on the ribosomesin the cytoplasm. Every protein must finally arrive in the part of the cell where it is to be functional. One of the central questions in molecular biology concerns the mechanism of protein sorting. The understanding of this issue is important for biotechnology, especially when it comes to direct recombinant proteins into the correct compartments.
Three important pathways of protein sorting ( Figure 5.1) are known:
Gated transport: Transport of proteins and RNA via the nuclear pore complex(NPC) into and out of the cell nucleus. The nuclear pores exhibit selective channeling, allowing entry only for certain macromolecules. The export out of the nucleus also proceeds selectively via nuclear pores.
Protein translocation: Uptake of a protein produced in the cytosol by an organelle via specific protein translocators. This is the pathway for proteins taken up by the mitochondria, plastids, and peroxisomes.
Vesicular transport: Proteins secreted in the endoplasmic reticulum(ER) undergo a series of posttranslational modifications in the ER and in the Golgi apparatus. The finished proteins are packed into vesicles and sent to the lysosomes, endosomes, or cytoplasmic membrane. There the vesicle fuses with the membranes of the organelles or the cell, and the content of the vesicle is released through exocytosis.
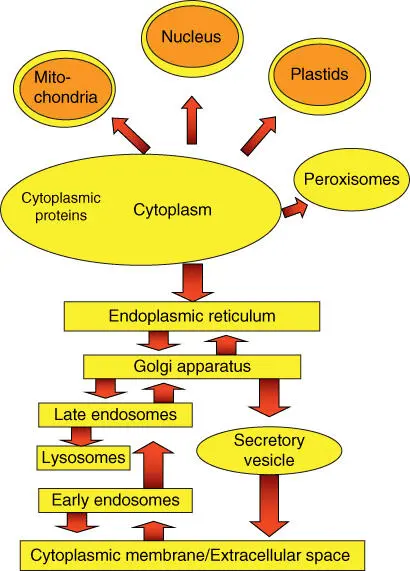
Figure 5.1 Schematic overview of protein transport inside a cell.
The selectivity of protein transportis based on recognition signalsthat proteins must carry. If a protein does not have a signal, it remains in the cytoplasm. All other proteins contain address labels that determine the designated location. They are either coherent signal sequences, with 15–60 amino acids, or signal patches, which are only recognizable in a three‐dimensional state and are made up of signal sequences from many protein domains. The signal sequences are very conserved in their structure. Important examples are shown in Table 5.1. Signal sequences are usually found on the N‐ or C‐terminal of a protein. They are usually removed by signal peptidasesas soon as a protein has reached its destination.
Table 5.1 Examples of typical recognition sequences.
| Targeted compartment | Sequence |
|---|---|
| Nuclear import | ‐Pro‐Pro‐ Lys‐Lys‐Lys‐Arg‐Lys‐Val‐ |
| Nuclear export | ‐ Met‐Glu‐Glu‐ Leu‐Ser‐Gln‐Ala‐Leu‐Ala‐Ser‐Ser‐ Phe‐ ‐ |
| Mitochondria | H 3N +‐Met‐Leu‐Ser‐Leu‐ Arg‐Gln‐Ser‐Ile‐ Arg‐Phe‐Phe‐ Lys‐Pro‐Ala‐Thr‐ |
| Arg‐Thr‐Leu‐Cys‐Ser‐Ser‐ Arg‐Tyr‐Leu‐Leu‐ | |
| Plastids | H 3N +‐Met‐Val‐Ala‐Met‐Ala‐Met‐Ala‐ Ser‐Leu‐Gln‐ Ser‐ Ser‐Met‐ Ser‐ Ser‐ |
| Leu‐ Ser‐Leu‐ Ser‐ Ser‐Asn‐ Ser‐Phe‐Leu‐Gly‐Gln‐Pro‐Leu‐ Ser‐Pro‐Ile‐ Thr‐ Leu‐ Ser‐Pro‐Phe‐Leu‐Gln‐Gly‐ | |
| Peroxisomes | ‐Ser‐Lys‐Leu‐COO – |
| ER import | H 3N +‐Met‐Met‐Ser‐Phe‐Val‐Ser‐ Leu‐Leu‐Leu‐Val‐Gly‐Ile‐Leu‐Phe‐Trp‐ |
| Ala‐Thr‐ Glu‐Ala‐Glu‐Gln‐Leu‐Thr‐ Lys‐Cys‐ Glu‐Val‐Phe‐Gln‐ | |
| ER retention | ‐ Lys‐Asp‐Glu‐Leu‐COO –‐ |
Amino acids printed in bold are especially important for the signal sequence.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «An Introduction to Molecular Biotechnology»
Представляем Вашему вниманию похожие книги на «An Introduction to Molecular Biotechnology» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.


![Andrew Radford - Linguistics An Introduction [Second Edition]](/books/397851/andrew-radford-linguistics-an-introduction-second-thumb.webp)









Обсуждение, отзывы о книге «An Introduction to Molecular Biotechnology» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.