Matthew B. Hamilton - Population Genetics
Здесь есть возможность читать онлайн «Matthew B. Hamilton - Population Genetics» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Population Genetics
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Population Genetics: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Population Genetics»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
is the classic, accessible introduction to the concepts of population genetics. Combining traditional conceptual approaches with classical hypotheses and debates, the book equips students to understand a wide array of empirical studies that are based on the first principles of population genetics.
Featuring a highly accessible introduction to coalescent theory, as well as covering the major conceptual advances in population genetics of the last two decades, the second edition now also includes end of chapter problem sets and revised coverage of recombination in the coalescent model, metapopulation extinction and recolonization, and the fixation index.
Population Genetics — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Population Genetics», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
The mixing of two genetically diverged populations, often termed admixture, can produce substantial levels of gametic disequilibrium. This is caused by different allele frequencies in the two source populations that result in different gamete frequencies at gametic equilibrium. Recombination acts to produce independent segregation but it does so only based on the allele frequencies within a group of mating individuals. Table 2.13gives an example of gametic disequilibrium produced when two populations with diverged allele frequencies are mixed equally to form a third population. In the example, the allele frequency divergence is large, and admixture produces a new population where gametic disequilibrium is 64% of its maximum value. In general, gametic disequilibrium due to the admixture of two diverged populations increases as allele frequencies become more diverged between the source populations, and the initial composition of the mixture population approaches equal proportions of the source populations.
Mating system
As covered earlier in this chapter, self‐fertilization and mating between relatives increase homozygosity at the expense of heterozygosity. An increase in homozygosity causes a reduction in the effective rate of recombination because crossing over between two homozygous loci does not alter the gamete haplotypes produced by that genotype. The effective recombination fraction under self‐fertilization is
(2.39) 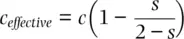
where s is the proportion of progeny produced by self‐fertilization each generation. This is based on the expected fixation index at equilibrium 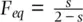 (Haldane 1924; see Figure 2.13). Figure 2.22shows the decay in gametic disequilibrium predicted by Eq. 2.39for four self‐fertilization rates in the cases of free recombination ( c = 0.5) and well as tight linkage ( c = 0.05). Self‐fertilization clearly increases the persistence of gametic disequilibrium, with marked effects at high selfing rates. In fact, the predominantly self‐fertilizing plant Arabidopsis thaliana exhibits gametic disequilibrium over much longer regions of chromosome compared to outcrossing plants and animals (see review by Flint‐Garcia et al. 2003, Kim et al. 2007).
(Haldane 1924; see Figure 2.13). Figure 2.22shows the decay in gametic disequilibrium predicted by Eq. 2.39for four self‐fertilization rates in the cases of free recombination ( c = 0.5) and well as tight linkage ( c = 0.05). Self‐fertilization clearly increases the persistence of gametic disequilibrium, with marked effects at high selfing rates. In fact, the predominantly self‐fertilizing plant Arabidopsis thaliana exhibits gametic disequilibrium over much longer regions of chromosome compared to outcrossing plants and animals (see review by Flint‐Garcia et al. 2003, Kim et al. 2007).
Table 2.13 Example of the effect of population admixture on gametic disequilibrium. In this case, the two populations are each at gametic equilibrium given their respective allele frequencies. When an equal number of gametes from each of these two genetically diverged populations are combined to form a new population, gametic disequilibrium results from the diverged gamete frequencies in the founding populations. The allele frequencies are: population 1 p 1= 0.1, p 2= 0.9, q 1= 0.1, q 2= 0.9; population 2 p 1= 0.9, p 2= 0.1, q 1= 0.9, q 2= 0.1. In population 1 and population 2, gamete frequencies are the product of their respective allele frequencies as expected under independent segregation. In the mixture population, all allele frequencies become the average of the two source populations (0.5) with D max= 0.25.
| Gamete | Gamete frequency | Population 1 | Population 2 | Mixture population | |
|---|---|---|---|---|---|
| A 1B 1 | g 11 | 0.01 | 0.81 | 0.41 | |
| A 2B 2 | g 22 | 0.81 | 0.01 | 0.41 | |
| A 1B 2 | g 12 | 0.09 | 0.09 | 0.09 | |
| A 2B 1 | g 21 | 0.09 | 0.09 | 0.09 | |
| D | 0.0 | 0.0 | 0.16 | ||
| D ' | 0.0 | 0.0 | 0.16/0.25 = 0.64 |

Figure 2.22 The decay of gametic disequilibrium ( D ) over time with random mating ( s = 0) and three levels of self‐fertilization. Initially, there are only coupling ( P 11= P 22= ½) and no repulsion gametes ( P 12= P 21= 0). Self‐fertilization slows the decay of gametic disequilibrium appreciably even when there is free recombination because double heterozygote genotypes are infrequent.
Population size
It is possible to observe gametic disequilibrium just by chance in small populations or small samples of gametes. Recombination itself is a random process in terms of where crossing over events occur in the genome. As shown in the Appendix, estimates are more likely to approach their true values as larger samples are taken. This applies to mating patterns and the number of gametes that contribute to surviving progeny in biological populations. If only a few individuals mate (even at random) or only a few gametes found the next generation, then this is a small “sample” of possible gametes that could deviate from independent segregation just by chance. When the chance effects due to population size and recombination are in equilibrium, the effects of population size can be summarized approximately by
(2.40) 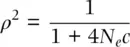
where N eis the genetic effective population size and c is the recombination fraction per generation (Hill and Robertson 1968; Ohta and Kimura 1969a, b; the basis of this type of equation is derived in Chapter 4). As shown in Figure 2.23, when the product of N eand c is small, chance sampling contributes to maintaining some gametic disequilibrium since only a few gametes contribute to the next generation when N eis small and genetic drift is strong, or only a few recombinant gametes exist when c is small. The lesson is that D as we have used it in this section assumes a large population size (similar to Hardy–Weinberg) so that actual gamete frequencies approach those expected based on allele frequencies, an assumption that is not met in actual populations to some degree because they are finite. Strong growth in population size over time can also alter the rate of decay of gametic disequilibrium compared to that seen in a population of constant size through time (Pritchard and Przeworski 2001; Rogers 2014).
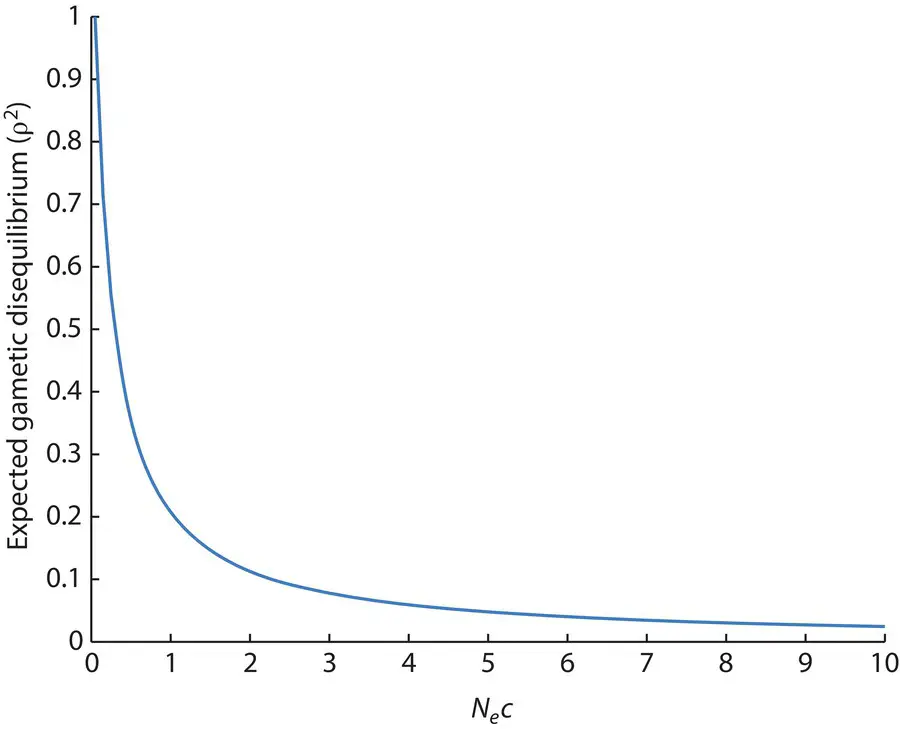
Figure 2.23 Expected levels of the squared gametic disequilibrium correlation (ρ 2) due to the combination of finite effective population size ( N e) and recombination at rate ( c ). Gametic disequilibrium is greater when fewer recombinant haplotypes are produced each generation (small c ), the population is small causing haplotype frequencies to fluctuate by chance (small N e), or if both factors are acting in combination (small N e c ).
Interact box 2.4 Estimating genotypic disequilibrium
In practice, the recombination fraction for two loci can be measured by crossing a double heterozygote with a double homozygote and then counting the recombinant gametes. However, this basic experiment cannot be carried out unless individuals can be mated in controlled crosses, excluding many, if not most, species. For two diploid loci, disequilibrium can occur between loci at two alleles positioned on the same chromosome, as well as between loci at two alleles positioned on different chromosomes (Weir 1996). With observed genotype data from pairs of loci where the phase of alleles or gamete organization is unknown, these latter two types of between‐locus disequilibrium cannot be distinguished but they can be considered together as genotypic disequilibrium (Rogers and Huff 2009).
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Population Genetics»
Представляем Вашему вниманию похожие книги на «Population Genetics» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Population Genetics» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.