Matthew B. Hamilton - Population Genetics
Здесь есть возможность читать онлайн «Matthew B. Hamilton - Population Genetics» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Population Genetics
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Population Genetics: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Population Genetics»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
is the classic, accessible introduction to the concepts of population genetics. Combining traditional conceptual approaches with classical hypotheses and debates, the book equips students to understand a wide array of empirical studies that are based on the first principles of population genetics.
Featuring a highly accessible introduction to coalescent theory, as well as covering the major conceptual advances in population genetics of the last two decades, the second edition now also includes end of chapter problem sets and revised coverage of recombination in the coalescent model, metapopulation extinction and recolonization, and the fixation index.
Population Genetics — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Population Genetics», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Linkage‐like effects can be seen in some chromosomes and genomes where gametic disequilibrium is expected to persist over longer time scales due to exceptional inheritance or recombination patterns. Organisms such as birds and primates have chromosomal sex determination, with the well‐known X and Y sex chromosome system in humans. Loci located on X chromosomes experience recombination, whereas those on Y chromosomes experience no recombination. This is caused by the Y chromosome lacking a homologous chromosome to pair with at meiosis since YY genotypes do not exist. In addition, we would expect that the rate of decay of gametic disequilibrium for X chromosomes is about half that of autosomes with comparable recombination rates, since X recombination takes place only in females (XX) at meiosis, and not at all in males (XY). Organelle genomes found in mitochondria and chloroplasts are a case where gametic disequilibrium persists indefinitely since these genomes are uniparentally inherited and do not experience observable levels of recombination. There is variation in the rate of recombination among species, within and among populations and between sexes (Stapley et al. 2017), with “hotspots” that show elevated rates as well as areas of restricted recombination such that genomes may have marked heterogeneity in recombination rates. In humans, for example, patterns of haplotype polymorphism suggest that about 80% of all recombination events take place in a subset of only about 15% of the genome (Myers et al. 2006).
Natural selection
Natural selection is a process that can continuously counteract the randomizing effects of recombination. Imagine a case where two locus genotypes confer different rates of survival or different levels of reproduction. In such a case, natural selection will reduce the frequency of lower fitness genotypes, which will also reduce the number of gametes these genotypes contribute to forming the next generation. At the same time that natural selection is acting, recombination is also working to randomize the associations of alleles at the two loci. Figure 2.21shows an example of epistatic natural selection acting to maintain gametic disequilibrium in opposition to recombination acting to establish gametic equilibrium. Natural selection at one locus can also impact the frequency of nearby loci that experience limited recombination. Both hitchhiking,where natural selection rapidly increasing the frequency of a beneficial genotype alters the frequency of linked loci, and background selection,where natural selection eliminates low fitness genotypes, can lead to linkage disequilibrium at loci adjacent to the loci experiencing selection. This is because natural selection has the potential to change the frequency of haplotypes more quickly than recombination can act to randomize the arrangement of alleles found together in the same haplotype. More on these natural selection topics can be found in later chapters.
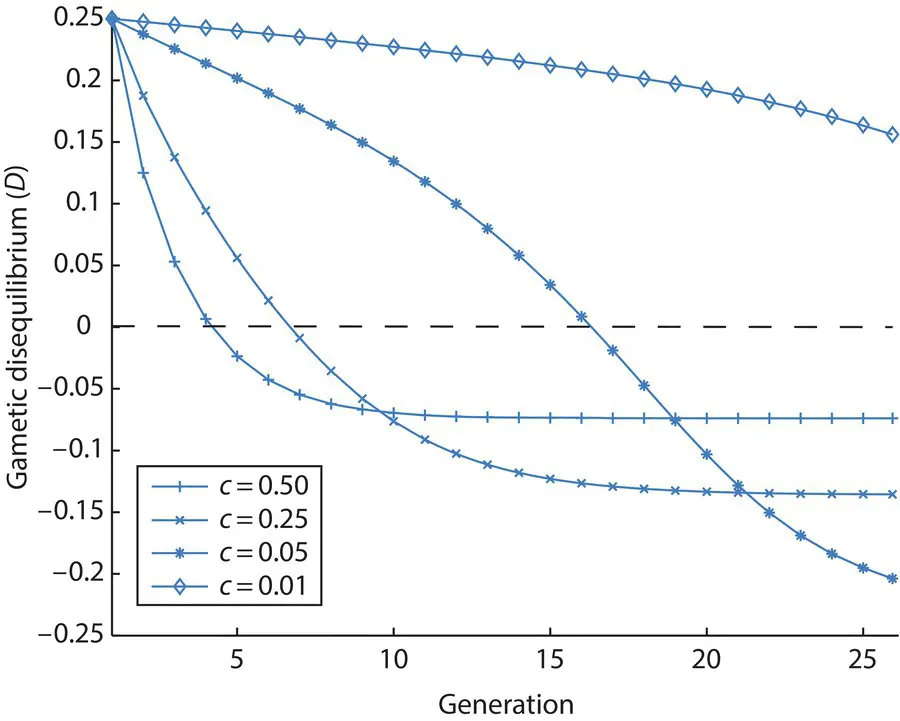
Figure 2.21 The decay of gametic disequilibrium ( D ) over time when both strong natural selection and recombination are acting. Initially, there are only coupling ( P 11= P 22= ½) and no repulsion gametes ( P 12= P 21= 0). The relative fitness values of the AAbb and aaBB genotypes are one, while all other genotypes have a fitness of 0.5, a form of epistasis for relative fitness. Unlike in Figure 2.19, gametic disequilibrium does not decay to zero over time due to natural selection that is stronger than recombination.
The action of natural selection acting on differences in gamete fitness can produce steady‐states other than D = 0 even with free recombination. In such cases, the population reaches a balance where the action of natural selection to increase the absolute value of D and the action of recombination to bring D back to zero cancel each other out. The point where the two processes are exactly equal in magnitude but opposite in their effects is where gametic disequilibrium will be maintained in a population. It is important to recognize that the amount of steady‐state gametic disequilibrium depends on which genotypes have high fitness values, so natural selection and recombination could also act in concert to accelerate the decay of gametic disequilibrium more rapidly than just recombination alone.
Interact box 2.3 Gametic disequilibrium under both recombination and natural selection
To simulate the combined action of recombination and natural selection on gametic disequilibrium, use the program Populus, which can be obtained by following the link on the text website.
In version 5.5 of Populus, use the Natural Selectionmenu to choose the Two‐Locus Selectionsimulation. Set p AB = p ab = 0.5 and p Ab = p aB = 0.0 as a case where there is maximum gametic disequilibrium initially. Use fitness values of w AaBb = 1, all others = 0.5 and w AAbb = w aaBB = 1, all others = 0.5 to generate strong natural selection via epistasis. Finally, try recombination values of r = 0.5 and 0.05. Focus your attention on the D vs. tplot. What do the two different fitness cases do to levels of gametic disequilibrium and how effective is recombination in opposing or accelerating this effect?
To see the details behind the recombination and natural selection model of gametic disequilibrium, a spreadsheet version of this model is available in Microsoft Excel format. The spreadsheet model will allow you to see all the calculations represented by formulas along with a graph of gametic disequilibrium over time. You can vary the recombination rate, initial gamete frequencies, and relative fitness values to see how they impact change in D and determine its eventual equilibrium.
Mutation
Alleles change from one form to another by the random process of mutation, which can either increase or decrease gametic disequilibrium. First consider the case of mutation producing a novel allele not found previously in the population. Since a new allele is present in the population as only a single copy, it is found only in association with the other alleles on the chromosome strand where it originated. Thus, a novel allele produced by mutation would initially increase gametic disequilibrium. Should the novel allele persist in the population and increase in frequency, then recombination will work to randomize the other alleles found with the novel allele and eventually dissipate the gametic disequilibrium. Mutation can also produce alleles identical to those currently present in a population. In that case, mutation can contribute to randomizing the combinations of alleles at different loci and thereby decrease levels of gametic disequilibrium. On the other hand, if the population is at gametic equilibrium mutation can create gametic disequilibrium by changing the frequencies of gamete haplotypes. However, it is important to recognize that mutation rates are often very low and the gamete frequency changes caused by mutation are inversely proportional to population size, so that mutation usually makes a modest contribution to overall levels of gametic disequilibrium. A simulation study showed that excluding any alleles at a frequency of less than 5–10% from estimates of D can eliminate most of the gametic disequilibrium attributable to recent mutations (Hudson 1985).
Mixing of diverged populations
Интервал:
Закладка:
Похожие книги на «Population Genetics»
Представляем Вашему вниманию похожие книги на «Population Genetics» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Population Genetics» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.