Matthew B. Hamilton - Population Genetics
Здесь есть возможность читать онлайн «Matthew B. Hamilton - Population Genetics» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Population Genetics
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Population Genetics: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Population Genetics»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
is the classic, accessible introduction to the concepts of population genetics. Combining traditional conceptual approaches with classical hypotheses and debates, the book equips students to understand a wide array of empirical studies that are based on the first principles of population genetics.
Featuring a highly accessible introduction to coalescent theory, as well as covering the major conceptual advances in population genetics of the last two decades, the second edition now also includes end of chapter problem sets and revised coverage of recombination in the coalescent model, metapopulation extinction and recolonization, and the fixation index.
Population Genetics — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Population Genetics», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Let's develop an example to understand the impact of mating among relatives on genotype and allele frequencies in a population. Under complete positive assortative mating or selfing, an individual mates with another individual possessing an identical genotype. Figure 2.12diagrams the process of positive genotypic assortative mating for a diallelic locus, following the frequencies of each genotype through time. Initially, the frequency of the heterozygote is H but this frequency will be halved each generation. A Punnett square for two heterozygotes shows that half of the progeny are heterozygotes ( H /2). The other half of the progeny are homozygotes ( H /2), composed of one‐quarter of the original heterozygote frequency of each homozygote genotype ( H /2[1–1/2]). It is obvious that matings among like homozygotes will produce only identical homozygotes, so the homozygote genotypes each yield a constant frequency of homozygous progeny each generation. In total, however, the frequency of the homozygous genotypes increases by a factor of 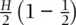 each generation due to homozygous progeny of the heterozygous genotypes. If the process of complete assortative mating continues, the population rapidly loses heterozygosity and approaches a state where the frequency of heterozygotes is zero.
each generation due to homozygous progeny of the heterozygous genotypes. If the process of complete assortative mating continues, the population rapidly loses heterozygosity and approaches a state where the frequency of heterozygotes is zero.
As an example, imagine a population where p = q = 0.5 that has Hardy–Weinberg genotype frequencies D = 0.25, H = 0.5, and R = 0.25. Under complete positive assortative mating, what would be the frequency of heterozygotes after five generations? Using Figure 2.12, at time t = 5, heterozygosity would be H (1/2) 5= H (1/32) = 1/64 or 0.016. This is a drastic reduction in only five generations.

Figure 2.12 The impact of complete positive genotypic assortative mating (like genotypes mate) or self‐fertilization on genotype frequencies. The initial genotype frequencies are represented by D , H , and R . When either of the homozygotes mates with an individual with the same genotype, all progeny bear their parent's homozygous genotype. When two heterozygote individuals mate, the expected genotype frequencies among the progeny are one half heterozygous genotypes and one quarter of each homozygous genotype. Every generation, the frequency of the heterozygotes declines by one half while one quarter of the heterozygote frequency is added to the frequencies of each homozygote (diagonal arrows). Eventually, the population will lose all heterozygosity, although allele frequencies will remain constant. Therefore, assortative mating or self‐fertilization changes the pairing of alleles in genotypes but not the allele frequencies themselves.
Genotype frequencies change quite rapidly under complete assortative mating, but what about allele frequencies? Let's employ the same example population with p = q = 0.5 and Hardy–Weinberg genotype frequencies D = 0.25, H = 0.5, and R = 0.25 to answer the question. For both of the homozygous genotypes, the initial frequencies would be D = R = (0.5) 2= 0.25. In Figure 2.12, the contribution of each homozygote genotype frequency from mating among heterozygotes after five generations is H /2(1 − (1/2) 5) = H /2(1–1/32) = H /2(31/32). With the initial frequency of H = 0.5, H /2(31/32) = 0.242. Therefore, the frequencies of both homozygous genotypes are 0.25 + 0.242 = 0.492 after five generations. It is also apparent that the total increase in homozygotes (31/32) is exactly the same as the total decrease in heterozygotes (31/32), so the allele frequencies in the population have remained constant. After five generations of assortative mating in this example, genotypes are much more likely to contain two identical alleles than they are to contain two unlike alleles. This conclusion is also reflected in the value of the fixation index for this example, 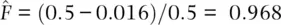 . In general, positive assortative mating or inbreeding changes the way in which alleles are “packaged” into genotypes , increasing the frequencies of all homozygous genotypes by the same total amount that heterozygosity is decreased, but allele frequencies in a population do not change.
. In general, positive assortative mating or inbreeding changes the way in which alleles are “packaged” into genotypes , increasing the frequencies of all homozygous genotypes by the same total amount that heterozygosity is decreased, but allele frequencies in a population do not change.
The fact that allele frequencies do not change over time can also be shown elegantly with some simple algebra. Using the notation in Figure 2.12and defining the frequency of the A allele as p and the a allele as q with subscripts to indicate generation, allele frequencies can be determined by the genotype counting method as 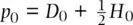 and
and 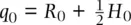 . Figure 2.12also provides the expressions for genotype frequencies from one generation to the next
. Figure 2.12also provides the expressions for genotype frequencies from one generation to the next 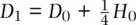 ,
, 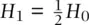 , and
, and 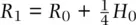 . We can then use these expressions to predict allele frequency in one generation:
. We can then use these expressions to predict allele frequency in one generation:
(2.14) 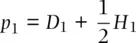
as a function of genotype frequencies in the previous generation using substitution for D 1and H 1:
(2.15) 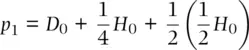
which simplifies to:
(2.16) 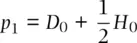
and then recognizing that the right‐hand side is equal to the frequency of A in generation 0:
(2.17) 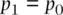
Thus, allele frequencies remain constant under complete assortative mating. As practice, you should carry out the algebra for the frequency of the a allele.
Under complete self‐fertilization heterozygosity declines very rapidly. There can also be partial self‐fertilization in a population (termed mixed mating), where some matings are self‐fertilization and others are between two individuals (called outcrossing). In addition, many organisms are not capable of self‐fertilization but instead engage in biparental inbreeding (mating between two different but related individuals) to some degree. In general, these forms of mating among relatives will reduce heterozygosity compared to random mating, although they will not drive heterozygosity toward zero as in the case of complete selfing. The rate of decline in heterozygosity can be determined for many possible types of mating systems, and a few examples are shown in Figure 2.13. Regardless of the specifics of the form of consanguineous mating that occurs, it remains true that mating among relatives causes alleles to be packaged more frequently as homozygotes (heterozygosity declines) and most forms of mating among relatives do not alter allele frequencies in a population. Negative assortative mating is an exception where allele frequencies can change depending on the initial allele frequencies (Workman 1964).
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Population Genetics»
Представляем Вашему вниманию похожие книги на «Population Genetics» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Population Genetics» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.