Matthew B. Hamilton - Population Genetics
Здесь есть возможность читать онлайн «Matthew B. Hamilton - Population Genetics» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Population Genetics
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Population Genetics: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Population Genetics»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
is the classic, accessible introduction to the concepts of population genetics. Combining traditional conceptual approaches with classical hypotheses and debates, the book equips students to understand a wide array of empirical studies that are based on the first principles of population genetics.
Featuring a highly accessible introduction to coalescent theory, as well as covering the major conceptual advances in population genetics of the last two decades, the second edition now also includes end of chapter problem sets and revised coverage of recombination in the coalescent model, metapopulation extinction and recolonization, and the fixation index.
Population Genetics — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Population Genetics», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Genetic genealogy or familial searching methods are used to identify suspects in unsolved serial violent crimes. While DNA profiles can be used as evidence to link a known suspect to a crime, direct identification of an unknown suspect is impossible if that person's DNA profile is not already present in a forensic or offender genetic database. A recent innovation is to use DNA profiles obtained from crime scenes to identify the relatives of suspects through searches in genetic genealogy databases. This approach initially utilized public databases to search for relatives and led to numerous cold cases being solved. At the same time, law enforcement and other uses of genealogical databases have generated concern over the privacy of genetic information.
(2.23) 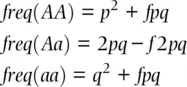
With consanguineous mating, the decline in realized heterozygosity (substituting H for freq[Aa] ) in the progeny is proportional to the coancestry coefficient which gives the probability that alleles identical in state are inherited because of relatedness among parents, shown by substituting H efor 2 pq in Eq. 2.20and rearranging to give
(2.24) 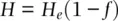
where H eis the Hardy–Weinberg expected heterozygosity based on population allele frequencies. Rearranging Eq. 2.21in terms of the autozygosity gives
(2.25) 
This is really exactly the same quantity as the fixation index ( Eq. 2.9)
(2.26) 
With the understanding that we are considering only a single generation of mating and that the population initially had Hardy–Weinberg expected genotype frequencies, we see that the fixation index measures an excess of homozygosity exactly equal to the autozygosity. Sustained non‐random mating over multiple generations will continue to increase the homozygosity, and therefore, F will reflect the cumulative deficit of heterozygosity from all past mating among relatives. In contrast, f is defined on a per generation basis and reflects only the probability of identity by descent at mating in the most recent generation. This distinction can be observed in Figure 2.13where F increases over time toward an equilibrium as a function of the coancestry coefficient due to each mating system.
We can also use the fixation index in Eq. 2.23rather than the autozygosity to express deviations from Hardy–Weinberg expected genotype frequencies due to accumulated autozygosity caused by mating patterns such as freq(AA) = p 2+ Fpq and freq(Aa) = 2 pq − F 2 pq where F 2 pq is the amount of heterozygosity missing from (or added to) the Hardy–Weinberg expected heterozygote frequency with half of that total or Fpq being added to (or subtracted from) each of the homozygotes. It is important to note that this time F represents the correlation of allelic state within genotypes from all causes accumulated over time that alters genotype frequencies from their Hardy–Weinberg expected values. While the difference in notation between F and f seems minor, their biological interpretations differ substantially in this example.
Returning to Figure 2.13reinforces the relationship of the coancestry coefficient, the fixation index, and the decline in heterozygosity in several specific cases of regular consanguineous mating. Remember that in all cases in Figure 2.13, the Hardy–Weinberg expected heterozygosity is 0.5 when mating is random and f = 0.
Phenotypic consequences of mating among relatives
The process of consanguineous mating is associated with changes in the mean phenotype within a population. These changes arise from two general causes: changes in genotype frequencies in a population per se and fitness effects associated with changes in genotype frequencies.
The mean phenotype of a population will be impacted by any changes in genotype frequency. To show this, it is necessary to introduce terminology to express the phenotype associated with a given genotype, a topic covered in much greater detail and explained more fully in Chapters 9and 10. We will assign AA genotypes the phenotype + a , heterozygotes the phenotype d , and aa homozygotes the phenotype − a . Each genotype contributes to the overall phenotype based on how frequent it is in the population. The mean phenotype in a population is then the sum of each genotype‐frequency‐weighted phenotype ( Table 2.10). When there is no dominance, the phenotype of the heterozygotes is exactly intermediate between the phenotypes of the two homozygotes and d = 0. In that case, it is easy to see that mating among relatives will not change the mean phenotype in the population since both homozygous genotypes increase by the same amount and their effects on the mean phenotype cancel out (mean = ap 2+ aFpq + d 2 pq − dF 2 pq − aq 2− aFpq , where the heterozygote terms are crossed out since d = 0). When there is some degree of dominance (positive d indicates the phenotype of Aa is like that of AA while negative d indicates the phenotype of Aa is like that of aa), then the mean phenotype of the population will change with consanguineous mating since heterozygotes will become less frequent. If dominance is in the direction of the + a phenotype ( d > 0), then mating among relatives will reduce the population mean because the heterozygote frequency will drop. Similarly, if dominance is in the direction of − a ( d < 0), then mating among relatives will increase the population mean again because the heterozygote frequency decreases. It is also true in the case of dominance that a return to random mating will restore the frequencies of heterozygotes and return the population mean to its original value mating among relatives. These changes in the population mean phenotype are simply a consequence of changing the genotype frequencies when there is no change in the allele frequencies.
There is a wealth of evidence that the increase of homozygosity caused by mating among relatives has deleterious(harmful or damaging) consequences and is associated with a decline in the average phenotype in a population, a phenomenon referred to as inbreeding depression. Since the early twentieth century, studies in animals and plants that have been intentionally inbred provide ample evidence that decreased performance, growth, reproduction, viability (all measures of fitness), and abnormal phenotypes are associated with consanguineous mating. A related phenomenon is heterosisor hybrid vigor, characterized by beneficial consequences of increased heterozygosity such as increased viability and reproduction, or the reverse of inbreeding depression. One example is the heterosis exhibited in corn, which has led to the widespread use of F1 hybrid seed in industrial agriculture.
Table 2.10 The mean phenotype in a population that is experiencing consanguineous mating. The fixation index quantifying deviation from Hardy–Weinberg expected genotype frequencies is F, and d = 0 when there is no dominance.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Population Genetics»
Представляем Вашему вниманию похожие книги на «Population Genetics» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Population Genetics» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.