Matthew B. Hamilton - Population Genetics
Здесь есть возможность читать онлайн «Matthew B. Hamilton - Population Genetics» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Population Genetics
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Population Genetics: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Population Genetics»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
is the classic, accessible introduction to the concepts of population genetics. Combining traditional conceptual approaches with classical hypotheses and debates, the book equips students to understand a wide array of empirical studies that are based on the first principles of population genetics.
Featuring a highly accessible introduction to coalescent theory, as well as covering the major conceptual advances in population genetics of the last two decades, the second edition now also includes end of chapter problem sets and revised coverage of recombination in the coalescent model, metapopulation extinction and recolonization, and the fixation index.
Population Genetics — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Population Genetics», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Once you are comfortable with the details of a simulation, you will also want to keep track of the “big picture” patterns that emerge as you view simulation results. Seeing these patterns will often require that you examine the results over a range of conditions. Try approaching simulations as experiments by changing only one variable at a time until you understand its effects on the outcome. Changing several things all at once can lead to confusion and an inability to see cause‐and‐effect relationships, unless you have fully understood the effects of individual variables. Finally, try writing down parameter values you have tried in a simulation and sketching or tabulating results on paper as you work with a simulation. Use all of your skills as a scientist and student when conducting simulations, and they will become a powerful learning tool. Eventually, you may even use scripting and programming to carry out your own simulations specifically designed to explore your own genetic hypotheses.
Chapter 1 review
Both general principles and direct measurements taken in actual populations combine to form comprehensive expectations about amounts, patterns, and cause‐and‐effect relationships in population genetics.
The theory of population genetics is the collection of well‐accepted expectations used to articulate a wide array of predictions about the biological processes that shape genetic variation.
Parameters are idealized quantities that are exact, while parameter estimates wear notational “hats” to remind us that they have statistical uncertainty.
Population genetics uses both inductive reasoning to generalize from the knowledge of specifics and deductive reasoning to build up predictions from general principles that can be applied to specific situations.
Population genetics is not a spectator sport! Direct participation through computer simulation provides the opportunity to see population genetic processes in action. You can learn by trial and error and test your own understanding by making predictions and then comparing them with simulation results.
Further reading
For a history of population genetics from Darwin to the 1930s, see:
1 Provine, W.B. (1971). The Origins of Theoretical Population Genetics. Chicago, IL: University of Chicago Press.For a concise history of population genetics since the mid‐1960s that highlights major conceptual advances as well as technical innovations to measure genetic variation, see:
2 Charlesworth, B. and Charlesworth, D. (2017). Population genetics from 1966 to 2016. Heredity 118: 2–9.For two personal and historical essays on the past, present, and assumptions of theoretical population genetics, see:
3 Lewontin, R.C. (1985). Population genetics. In: Evolution: Essays in Honour of John Maynard Smith (eds. P.J. Greenwood, P.H. Harvey and M. Slatkin), 3–18. Cambridge: Cambridge University Press.
4 Wakeley, J. (2005). The limits of theoretical population genetics. Genetics 169: 1–7.
CHAPTER 2 Genotype frequencies
2.1 Mendel's model of particulate genetics
Mendel's breeding experiments.
Independent assortment of alleles.
Independent segregation of loci.
Some common genetic terminology.
In the nineteenth century, there were several theories of heredity, including inheritance of acquired characteristics and blending inheritance. Jean‐Baptiste Lamarck is most commonly associated with the discredited hypothesis of inheritance of acquired characteristics (although it is important to recognize his efforts in seeking general causal explanations of evolutionary change). He argued that individuals contain “nervous fluid” and that organs or features (phenotypes) employed or exercised more frequently attract more nervous fluid, causing the trait to become more developed in their offspring. His widely known example is the long neck of the giraffe, which he said developed because individuals continually stretched to reach leaves at the tops of trees. Later, Charles Darwin and many of his contemporaries subscribed to the idea of blending inheritance. Under blending inheritance, offspring display phenotypes that are an intermediate combination of parental phenotypes ( Figure 2.1).
From 1856 to 1863, the Augustinian monk Gregor Mendel carried out experiments with pea plants that demonstrated the concept of particulate inheritance. Mendel showed that phenotypes are determined by discrete units that are inherited intact and unchanged through generations. His hypothesis was sufficient to explain three common observations: (i) phenotype is sometimes identical between parents and offspring; (ii) offspring phenotype can differ from that of the parents; and (iii) “pure” phenotypes of earlier generations could skip generations and reappear in later generations. Neither blending inheritance nor inheritance of acquired characteristics are satisfactory explanations for all of these observations. It is hard for us to fully appreciate now, but Mendel's results were truly revolutionary and served as the very foundation of population genetics. The lack of an accurate mechanistic model of heredity severely constrained biological explanations of cause and effect up to the point that Mendel's results were “rediscovered” in the year 1900.
It is worthwhile to briefly review the experiments with pea plants that Mendel used to demonstrate independent assortment of both alleles within a locus and of multiple loci, sometimes dubbed Mendel's first and second laws. We need to remember that this was well before the Punnett square, which originated in about 1905. Therefore, the conceptual tool we would use now to predict progeny genotypes from parental genotypes was a thing of the future. So, in revisiting Mendel's experiments, we will not use the Punnett square in an attempt to follow his logic. Mendel only observed the phenotypes of generations of pea plants that he had hand‐pollinated. From these phenotypes and their patterns of inheritance, he inferred the existence of heritable factors. His experiments were actually both logical and clever, but are now taken for granted since the basic mechanism of particulate inheritance has long since ceased to be an open question. It was Mendel who established the first and most fundamental prediction of population genetics: expected genotype frequencies.
Mendel used pea seed coat color as a phenotype he could track across generations. His goal was to determine, if possible, the general rules governing the inheritance of pea phenotypes. He established “pure”‐breeding lines (meaning plants that always produced progeny with phenotypes like themselves) of peas with both yellow and green seeds. Using these pure‐breeding lines as parents, he crossed a yellow‐ and a green‐seeded plant. The parental cross and the next two generations of the progeny are shown in Figure 2.2. Mendel recognized that the F1 plants had an “impure” phenotype because of the F2 generation plants, of which three‐quarters had yellow and one‐quarter had green seed coats.
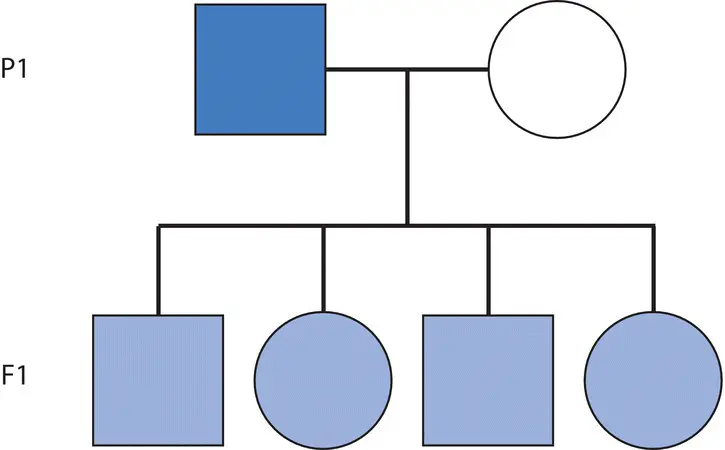
Figure 2.1 The model of blending inheritance predicts that progeny have phenotypes that are the intermediate of their parents. Here, “pure” blue and white parents yield light blue progeny, but these intermediate progeny could never themselves be parents of progeny with pure blue or white phenotypes identical to those in the P1 generation. Crossing any shade of blue with a pure white or blue phenotype would always lead to some intermediate shade of blue. By convention, in pedigrees, females are indicated by circles and males by squares while “P” refers to parental and “F” to filial.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Population Genetics»
Представляем Вашему вниманию похожие книги на «Population Genetics» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Population Genetics» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.