Luis A. Ebensperger - Comportamiento social de la fauna nativa de Chile
Здесь есть возможность читать онлайн «Luis A. Ebensperger - Comportamiento social de la fauna nativa de Chile» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на испанском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Comportamiento social de la fauna nativa de Chile
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Comportamiento social de la fauna nativa de Chile: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Comportamiento social de la fauna nativa de Chile»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
El comportamiento social ha enfatizado el uso de unas pocas especies de laboratorio, así como especies silvestres de regiones distintas de Chile y otras regiones vecinas. Este libro explora los estudios realizados en especies menos conocidas, como algunos vertebrados e invertebrados nativos de Chile, los cuales tienen un alto potencial para contribuir directamente a un desarrollo integrativo de la teoría existente.
En los capítulos de este libro se aborda la diversidad de elementos del comportamiento social examinados y cómo dichos elementos difieren o se asemejan a aquellos de otras especies ya estudiadas. Además, se entrega organizadamente información hasta ahora dispersa sobre variados atributos del comportamiento social de especies nativas de Chile. Se enfatiza la necesidad e importancia de desarrollar aún más el estudio científico del comportamiento social en especies nativas.
El presente volumen busca educar a un público no especializado sobre la singularidad y el valor científico del comportamiento social que caracteriza a la fauna nativa de este país. Además, se espera incentivar a estudiantes en formación e investigadores de disciplinas afines a considerar modelos sociales locales y explorar nuevos organismos de estudio para abordar problemáticas novedosas e integrativas.
Comportamiento social de la fauna nativa de Chile — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Comportamiento social de la fauna nativa de Chile», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:

Grupos sociales: roedores
Las descripciones de la organización y estructura social de roedores típicamente se basan en la captura, marcaje individual, seguimiento (ej., mediante telemetría), y registro de interacciones sociales a través de observación directa. Esta información ha permitido determinar el número y composición individual de grupos sociales distintivos, así como la naturaleza de las relaciones sociales predominantes entre sus integrantes, en 20 especies de roedores con distribución en Chile (Tabla 3-2). Nueve de estas especies excavan y utilizan comunalmente madrigueras como refugio. La organización social en estas últimas especies exhibe variabilidad intrapoblacional, y donde la diversidad de unidades (tipos) sociales registradas varía entre dos ( Aconaemys porteri ) y siete ( Octodon degus ) (Tabla 3-2). La existencia de grupos sociales distintivos también ha sido confirmada en dos poblaciones de cururos en Chile, una en la cordillera de los Andes de Chile central (La Parva) y otra en la costa del centro-norte de Chile (Parque Nacional Bosque Fray Jorge). Esta especie además es colonial y los individuos se alimentan de estructuras subterráneas que estos animales alcanzan a través de expandir activamente los túneles de sus galerías (Reig 1970, Begall y Gallardo 2000).
En degus ( O. degus ), la variabilidad social incluye hembras y machos solitarios, cohabitación estable entre parejas hembra-macho, y grupos con varios individuos de ambos sexos (Tabla 3-2, Figura 3-10a). El tamaño de los grupos varía entre poblaciones y años (Ebensperger et al . 2012, Sobrero et al . 2016a). Por otra parte, los grupos sociales de S. cyanus en la localidad de La Parva muestran una distribución más dispersa y menor cohesión social dentro de los grupos comparado con aquellos estudiados en el Parque Nacional Bosque Fray Jorge (Lacey et al . 2019). La organización social también es variable entre poblaciones ecológicamente divergentes en el cui de la Patagonia ( Microcavia australis ) en Argentina (Taraborelli y Moreno 2009, Correa Capítulo 5). En estas tres especies la variabilidad social reportada covaría con las condiciones ecológicas (Taraborelli y Moreno 2009, Ebensperger et al . 2012, Lacey et al . 2019). En cambio, la organización social registrada en socos ( Octodontomys gliroides , Figura 3-10b) es relativamente invariable aún en poblaciones con condiciones ecológicas contrastantes a ambos lados de Los Andes entre Chile y Bolivia (Rivera et al . 2014, Correa Capítulo 5).
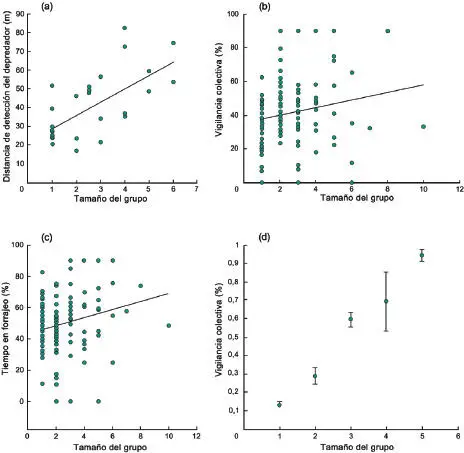
La variabilidad (o constancia) social registrada en degus, socos y cuyes de la Patagonia ha permitido realizar inferencias sobre sus posibles causas. Estudios en degus y cuyes apoyan la ocurrencia de beneficios sociales derivados de una disminución en el riesgo de depredación. Degus que forrajean en grupos más numerosos son más eficientes en detectar posibles depredadores (Figura 3-11a), a través de incrementar su capacidad de vigilancia colectiva (Figura 3-11b)(Ebensperger et al . 2006a, Ebensperger y Wallem 2002), lo que les permite dedicar más tiempo a forrajear (Vásquez 1997, Ebensperger et al . 2006a, Figura 3-11c). Una reducción social del riesgo de depredación también es consistente con la observación en la que degus forman grupos más numerosos en poblaciones y años con mayor abundancia de depredadores y menor densidad de refugios (Ebensperger et al . 2012). Es interesante que estos animales vigilan más frecuentemente utilizando una postura bípeda (Figura 3-10a)en parches con vegetación herbácea más alta, y visualmente más obstructiva (Ebensperger et al . 2005). Dado que la adopción de una postura bípeda podría ser más costosa en términos de disminuir la eficiencia para localizar alimento a nivel del suelo, también sería esperable un efecto social sobre el tipo de vigilancia en esta y otras especies. El riesgo de depredación también afecta el comportamiento de vigilancia del cuis durante el forrajeo social, y donde individuos en grupos más numerosos aumentan su vigilancia colectiva (Figura 3-11d), lo que presumiblemente aumenta su capacidad para detectar depredadores (Taraborelli 2008). De igual modo, el tamaño de los grupos en esta especie es mayor en poblaciones con menor cobertura arbustiva (i.e., refugio contra depredadores aéreos) y temperaturas más bajas, lo que además es consistente con beneficios energéticos derivados de termorregular socialmente (Taraborelli y Moreno 2009).
Figura 3-10 a)Degus ( Octodon degus ) de un mismo grupo social realizando vigilancia en posición bípeda luego de una vocalización de alarma en Rinconada de Maipú, Chile central; b)socos ( Octodontomys gliroides ) de un mismo grupo social en contacto físico en Chusmiza (norte de Chile) durante el atardecer, posiblemente para termorregular socialmente. Imágenes gentileza de (a) Juan Riquelme, y (b) Daniela Rivera.

Figura 3-11 a)Degus ( Octodon degus ) en grupos más numerosos son más eficientes para detectar la aproximación de potenciales depredadores, b)presumiblemente a través de incrementar su vigilancia colectiva, c)lo que les permite destinar más tiempo a forrajeo; (modificadas de Ebensperger et al . 2006a, Ebensperger y Wallem 2002); d)cuises ( Microcavia australis ) en grupos más numerosos también incrementan su vigilancia colectiva (modificada de Taraborelli 2008).
Por otra parte, el posible efecto de limitaciones en la disponibilidad de alimento parece menos importante en degus y socos. En degus la abundancia de alimento no cambia con la cercanía a madrigueras utilizadas por grupos de distinto tamaño (Hayes et al . 2009). Sin embargo, la abundancia de alimento es espacialmente más heterogénea en áreas de forrajeo utilizadas por grupos más numerosos (Ebensperger et al . 2016a). Un estudio preliminar indicó además que ni la cantidad ni la calidad (en base a la abundancia de alimento) de las madrigueras utilizadas covarían con la densidad de degus, algo que no apoyaría la ocurrencia de restricciones basadas en estos recursos (Ebensperger et al . 2011). Así, el posible efecto social de la distribución de alimento parece ser relativamente limitado en degus. De igual modo, el tamaño de los grupos sociales en socos tampoco covaría con diferencias en la abundancia y heterogeneidad de alimento y refugio entre poblacionales (Rivera et al . 2014).
Todas las especies de roedores nativos de Chile donde se ha registrado vida en grupos sociales corresponden a Histricomorfos del Nuevo Mundo (o caviomorfos), un grupo filogenéticamente hermano de los Phiomorfos del Viejo Mundo, que incluye a las ratas africanas (Upham y Patterson 2012). Análisis filogenéticos indican que la sociabilidad en caviomorfos ha estado asociada al riesgo de depredación, así como a la construcción y uso comunal de madrigueras subterráneas. Especies donde los grupos sociales son más numerosos también son frecuentemente más diurnas, con individuos de mayor tamaño corporal (i.e., más conspicuas a depredadores visuales diurnos), y que excavan madrigueras activamente (Ebensperger y Cofré 2001, Ebensperger y Blumstein 2006). Por otra parte, la sociabilidad en caviomorfos y phiomorfos es un rasgo ancestral, donde la reversión evolutiva de este rasgo a vida solitaria ha estado asociada al uso de hábitats con menor cobertura arbustiva y arbórea, algo que también apoya un efecto del riesgo de depredación como causa de la vida en grupos sociales (Sobrero et al . 2014b). Sin embargo, la evolución de la sociabilidad en estos organismos también ha estado influenciada por factores históricos. Específicamente, la sociabilidad en estos roedores es en parte consecuencia de la sociabilidad que caracterizó a sus ancestros. De este modo, la historia evolutiva de estos organismos ha estado marcada por algún grado de desacople entre la ocurrencia vida social y las condiciones ecológicas de los ambientes utilizados por estos (Rivera et al . 2014, Sobrero et al . 2014b).
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Comportamiento social de la fauna nativa de Chile»
Представляем Вашему вниманию похожие книги на «Comportamiento social de la fauna nativa de Chile» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Comportamiento social de la fauna nativa de Chile» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.