Rudy Bilous - Handbook of Diabetes
Здесь есть возможность читать онлайн «Rudy Bilous - Handbook of Diabetes» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Handbook of Diabetes
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Handbook of Diabetes: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Handbook of Diabetes»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Now in its fifth edition, this market-leading book aligns with the most recent guidelines from the American Diabetes Association (ADA), the European Association for the Study of Diabetes (EASD), Diabetes UK, and the National Institute for Health and Care Excellence (NICE), presenting authoritative clinical coverage of diabetes in an accessible format with rich pedagogical features.
Five new chapters provide detailed coverage of liver disease, diabetes education, bariatric surgery, diabetes and cancer, and the use of incretin-based therapies and SGLT2 Inhibitors in the management of Type II diabetes Updated and expanded topics include the relation between hypoglycaemia and dementia, anxiety and depression, the NICE Quality and Outcomes Framework (QOF), and the impacts of diabetes to self-care, mental health, and decision-making Provides a wealth of pedagogical features such as vignettes and case histories, important learning points, summaries of key clinical trials, and links to further readings
remains the essential practical companion for all health professionals involved in managing patients with diabetes, and an up-to-date reference for diabetes and endocrinology researchers, scientists, and academics.
Handbook of Diabetes — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Handbook of Diabetes», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
The main cell types of the pancreatic islets are β‐cells that produce insulin, α‐cells that secrete glucagon, δ‐cells that produce somatostatin and PP‐cells that produce pancreatic polypeptide. Different islet cell types can be identified by various immunostaining techniques. β‐cells are the most numerous cell type and are located mainly in the core of the islet, while α and δ cells are located in the periphery ( Figure 5.2).
Islet cells interact with each other through direct physical contact and through paracrine effects of their hormone products (e.g. glucagon stimulates insulin secretion and somatostatin inhibits insulin and glucagon secretion) ( Figure 5.3). The blood flow within the islets is organised centrifugally so that the different cell types are supplied in the sequence β → α → δ. Insulin also has an ‘autocrine’ (self‐regulating) effect that alters the transcription of insulin and glucokinase genes in the β cell.
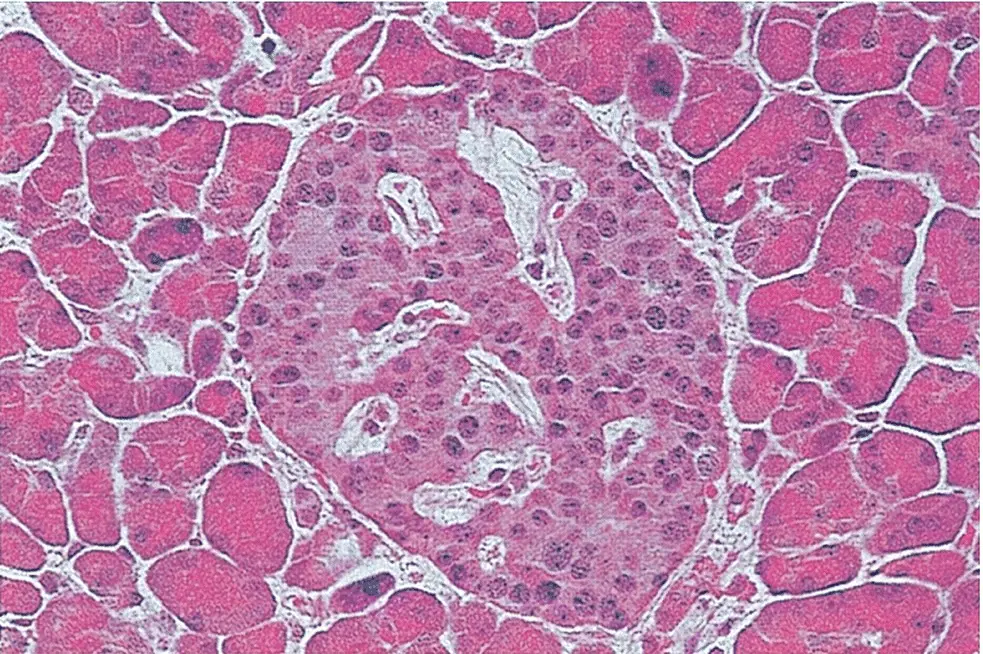
Figure 5.1 A section of normal pancreas stained with haematoxylin and eosin. As observed by Paul Langerhans, the islet in the centre is identified easily by its distinct morphology and lighter staining than that of the surrounding exocrine tissue (original magnification ×350).

Figure 5.2 The localisation of pancreatic hormones in human islets. (a) Insulin immunostained in the majority of cells that form the core of the islet (peroxidase–antiperoxidase immunostain with haematoxylin counterstain). (b) Insulin mRNA localised by in situ hybridization with a digoxigenin‐labelled sequence of rat insulin cRNA (which cross‐reacts fully with human insulin mRNA). (c) Peripherally located α cells immunostained with antibodies to pancreatic glucagon using the same method as for (a). (d) Weakly immunoreactive PP cells in the epithelium of a duct in the ventral portion of the pancreatic head. Magnifications approximately ×150.
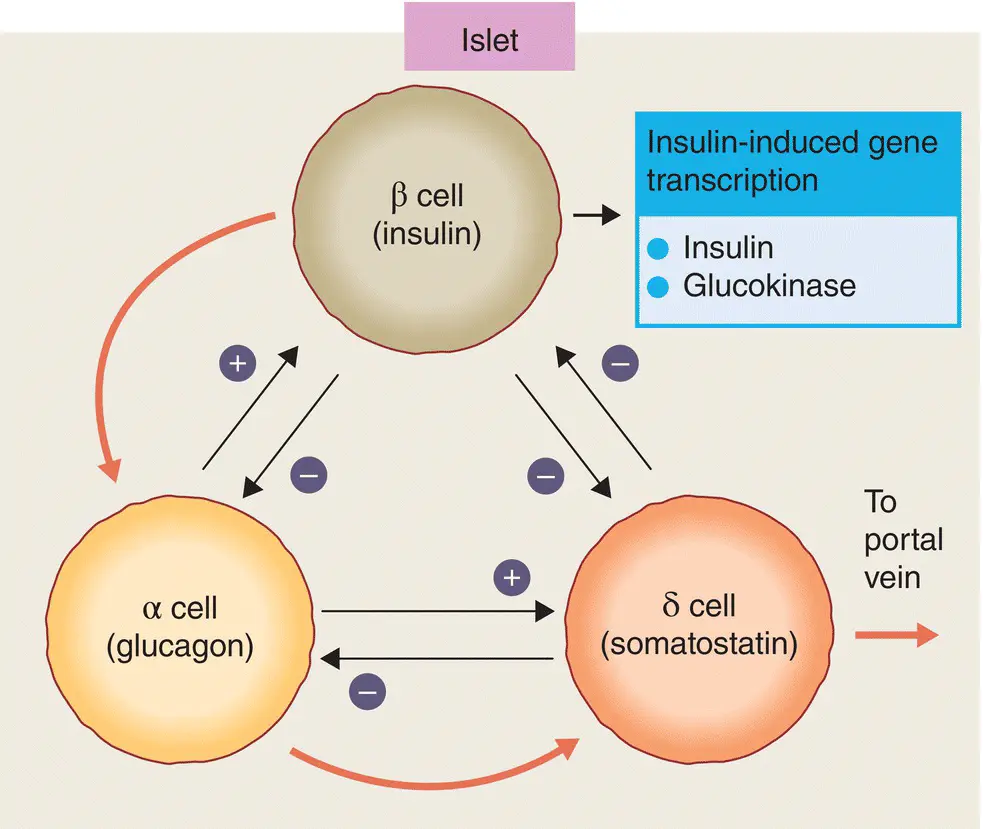
Figure 5.3 Potential interactions between the secretory products of the major islet cell types. Black arrows indicate paracrine stimulation or inhibition. The direction of blood flow within the islet is indicated by the red arrows.
Pancreatic islets are densely innervated with autonomic and peptidergic nerve fibres ( Figure 5.4). Parasympathetic innervation from the vagus stimulates insulin release, while adrenergic sympathetic nerves inhibit insulin and stimulate glucagon secretion. Other nerve fibres that supply the pancreas release peptides which also regulate pancreatic function, e.g vasoactive intestinal peptide (VIP) stimulates the release of all islet hormones and neuropeptide Y (NPY) inhibits insulin secretion. Neural pathways are important in the regulation of pancreatic function.
Pancreatic β‐cells may change in size, number, and function during normal aging and development ( Figure 5.5). β cell mass is determined by the net effect of four independent mechanisms: (i) β cell replication (i.e. division of existing β cells), (ii) β cell size, (iii) β cell neogenesis (i.e. emergence of new β cells from pancreatic ductal epithelial cells), and (iv) β cell apoptosis. The contribution made by each of these processes is variable and may change at different stages of life.
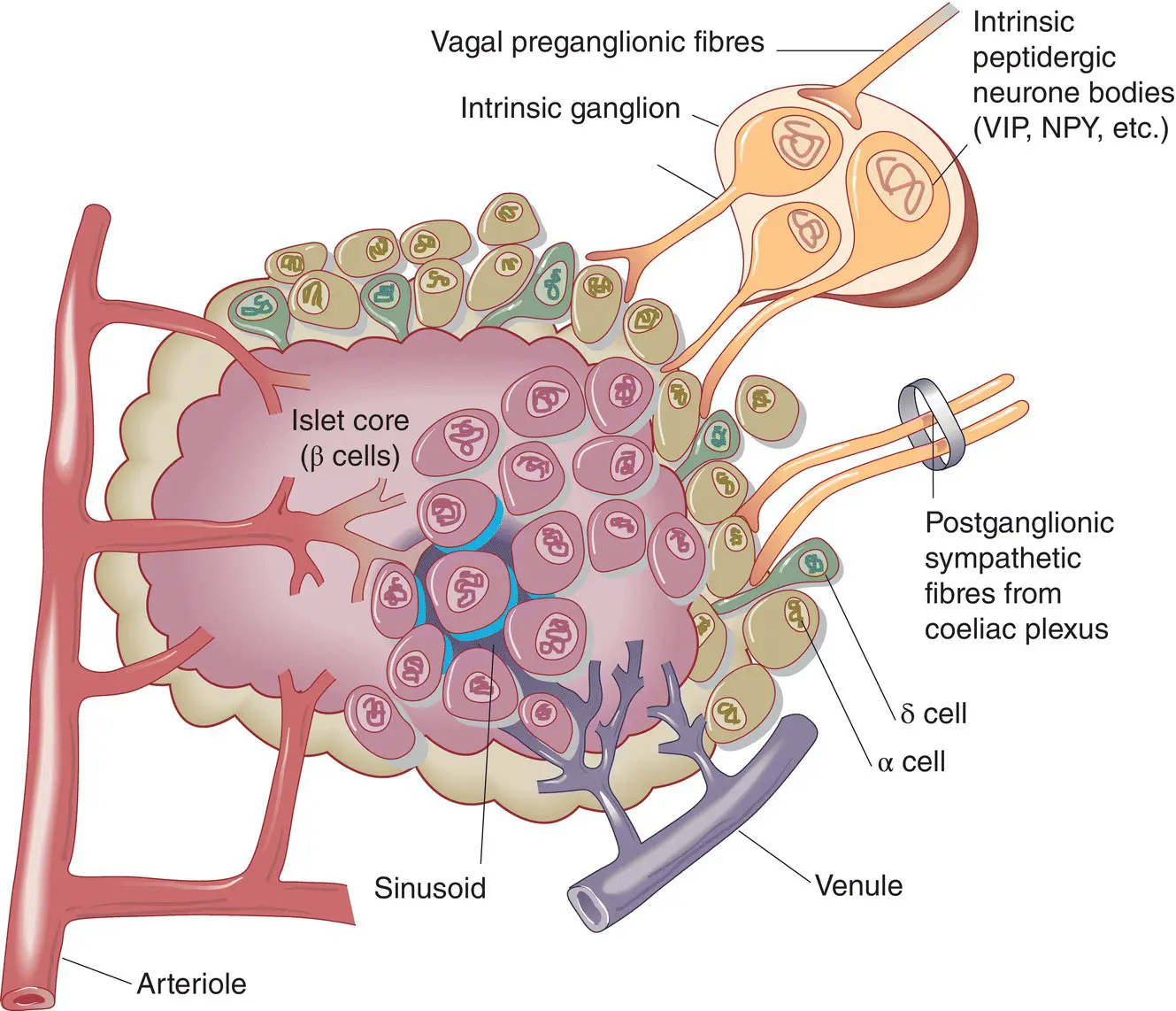
Figure 5.4 Structure of a pancreatic islet, showing the anatomical relationships between the four major endocrine cell types. NPY, neuropeptide Y; VIP, vasoactive intestinal polypeptide.
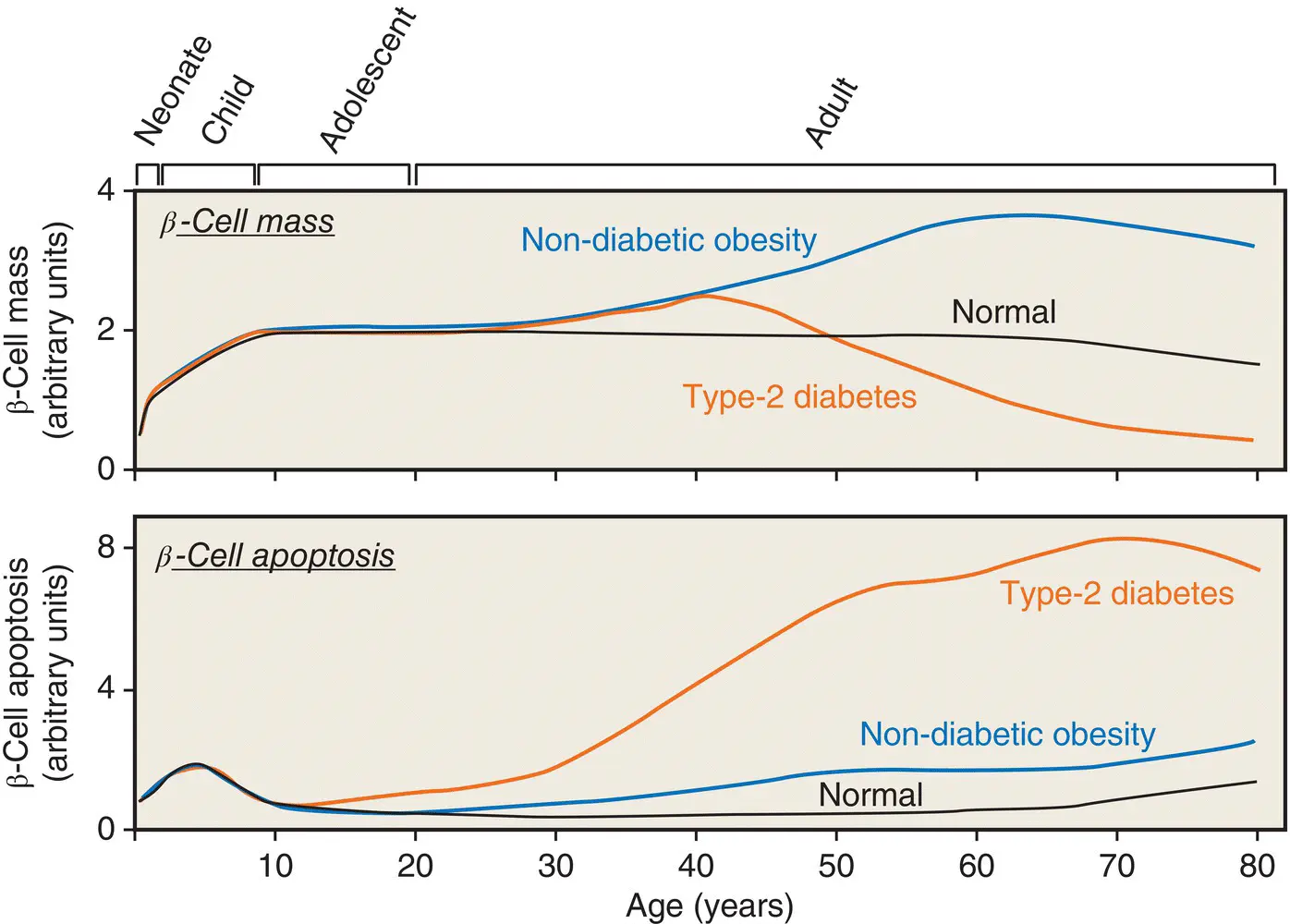
Figure 5.5 A hypothetical model for postnatal pancreatic β cell growth in humans.
Adapted from Rhodes et al. Science 2005; 307: 380–384.
Insulin synthesis and insulin polypeptide structure
Insulin consists of two polypeptide chains linked by disulphide bridges; the A‐chain contains 21 amino acids and the B‐chain contains 30 amino acids ( Figure 5.6). In the circulation, insulin exists as a monomer of 6000 Da molecular weight. The tertiary (three‐dimensional) structure of monomeric insulin consists of a hydrophobic core buried beneath a surface that is hydrophilic, except for two non‐polar regions involved in the aggregation of the monomers into dimers and hexamers.
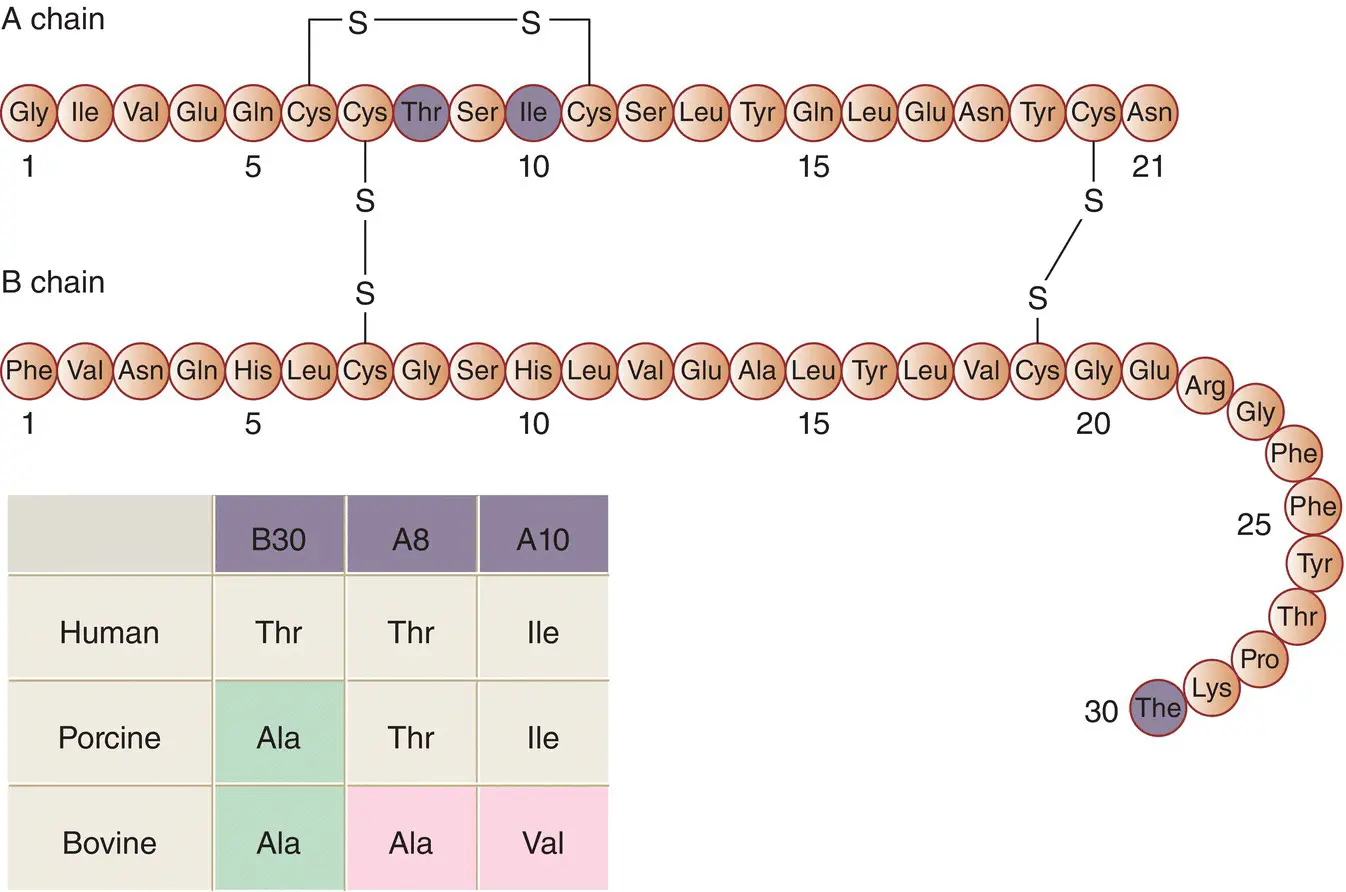
Figure 5.6 The primary structure (amino acid sequence) of human insulin. The highlighted residues are those that differ in porcine and bovine insulins, as shown in the inset.
In concentrated solution (such as in the insulin vial supplied by the pharmaceutical company for injection) and in crystals (such as in the insulin secretory granule), six monomers self‐associate with two zinc ions to form a hexamer ( Figure 5.7). This is of therapeutic importance because the slow absorption of native insulin from the subcutaneous tissue partly results from the time taken for the hexameric insulin to dissociate into the smaller, more easily absorbed monomeric form.
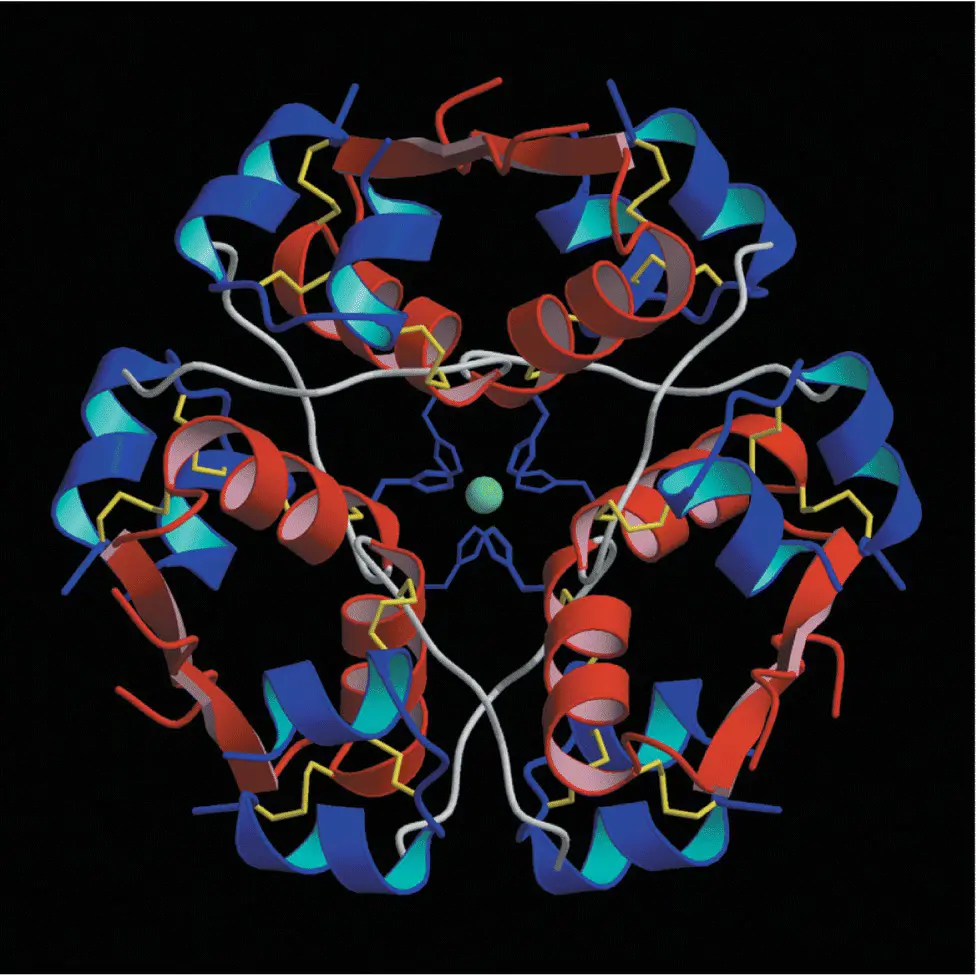
Figure 5.7 The double zinc insulin hexamer composed of three insulin dimers in a threefold symmetrical pattern.
Insulin is synthesised in the β cells from a single amino acid precursor called proinsulin ( Figure 5.8). Synthesis begins with the formation of an even larger precursor, preproinsulin, which is cleaved by protease activity to proinsulin. The gene for preproinsulin (and therefore the ‘gene for insulin’) is located on chromosome 11. Proinsulin is packaged into vesicles in the Golgi apparatus of the β cell; in the maturing secretory granules that bud off it, proinsulin is converted by enzymes into insulin and connecting peptide (C‐peptide).
Insulin and C‐peptide are released from the β cell when the secretory granules are transported (‘translocated’) to the cell surface and fuse with the plasma membrane (exocytosis) ( Figure 5.9). Microtubules, formed of polymerised tubulin, probably provide the mechanical framework for granule transport, and microfilaments of actin, interacting with myosin and other motor proteins such as kinesin, may provide the mechanical force that propels the granules along the tubules. Although the actin cytoskeleton is a key mediator of biphasic insulin release, cyclic GTPases are involved in F‐actin reorganization in the islet β cell and play a crucial role in stimulus‐secretion coupling.
The ‘regulated pathway’, with almost complete cleavage of proinsulin to insulin, normally accounts for about 95% of the β cell insulin production. In certain conditions, however, e.g insulinoma and type 2 diabetes, an alternative ‘constitutive’ pathway operates, in which large amounts of unprocessed proinsulin and intermediate insulin precursors (‘split proinsulins’) are released directly from vesicles that originate in the endoplasmic reticulum ( Figure 5.10).
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Handbook of Diabetes»
Представляем Вашему вниманию похожие книги на «Handbook of Diabetes» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Handbook of Diabetes» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.