Keith N. Frayn - Human Metabolism
Здесь есть возможность читать онлайн «Keith N. Frayn - Human Metabolism» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Human Metabolism
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Human Metabolism: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Human Metabolism»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
(formerly
) offers a current and integrated review of metabolism and metabolic regulation. The authors explain difficult concepts in clear and concise terms in order to provide an accessible and essential guide to the topic. This comprehensive text covers a wide range of topics such as energy balance, body weight regulation, exercise, and how the body copes with extreme situations, and illustrates how metabolic regulation allows the human body to adapt to many different conditions.
This fourth edition has been revised with a new full colour text design and helpful illustrations that illuminate the regulatory mechanisms by which all cells control the metabolic processes necessary for life. The text includes chapter summaries and additional explanatory text that help to clarify the information presented. In addition, the newly revised edition includes more content on metabolic pathways and metabolic diseases. This important resource:
Is a valuable tool for scientists, practitioners and students across a broad range of health sciences including medicine, biochemistry, nutrition, dietetics, sports science and nursing Includes a full colour text filled with illustrations and additional diagrams to aid understanding Offers a companion website with additional learning and teaching resources. Written for students of medicine, biochemistry, nutrition, dietetics, sports science and nursing,
has been revised and updated to provide a comprehensive review of metabolism and metabolic regulation.
Human Metabolism — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Human Metabolism», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
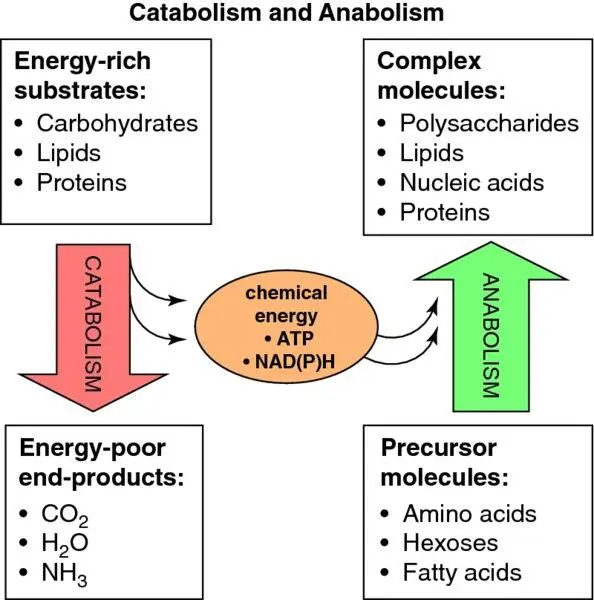
Figure 1.12 Catabolism and anabolism.
Box 1.4 Anabolism and catabolism
The terms anabolism and catabolism are useful but can be confusing and have frequently been misused. They should be used to refer to whole-bodyenergy strategy:

Hence, in the postprandial state, after a meal, we are entering an anabolic state, whereas in the post-absorptive state, following absorption and disposition of the meal, we are entering a catabolic state. This is signalled by insulin.
Classic physiological catabolic states include fasting/starvation (decreased energy intake) and exercise (increased energy expenditure). Diabetes mellitus is an example of a pathological catabolic state (failure of insulin signalling).
If the terms are applied to individual metabolic pathways, or even individual steps, confusion can arise. For example glycolysis may be thought of as ‘catabolic’ in exercising muscle, breaking down glucose to provide energy for contraction (net energy mobilisation), but ‘anabolic’ in liver in the well-fed postprandial state, when absorbed glucose is converted to pyruvate, but the resulting acetyl-CoA undergoes lipogenesis to fat for energy storage. When analysing metabolism it is important to consider the whole body (anabolic? catabolic?) as well as individual tissues, as these all have specialised metabolic profiles and functions (see Chapter 5).
The body is subject to many catabolic signals (e.g. ‘stress hormones,’ catecholamines, glucocorticoids, glucagon etc., but one major anabolic signal – insulin. Insulin inhibits catabolism, and therefore when it declines, unopposed catabolism results. This is one rea- son why insulin is such a crucial signal, and diabetes such an important disease.
The rate of energy production is measured under basal conditions (no voluntary muscle contraction; thermoneutrality) – ‘basal metabolic rate’ (BMR), and is affected by many factors, including muscle contraction, food ingestion, size, gender, age, temperature, sepsis, and several hormones, including thyroid hormones and catecholamines. The metabolic rate can be estimated by measuring the oxygen consumption (VO 2; indirect calorimetry). For carbohydrate metabolism the rate of CO 2production (VCO 2) equals VO 2(C 6H 12O 6+ 6O 2→ 6CO 2+ 6H 2O) and the ratio VCO 2/VO 2, termed the respiratory quotient (RQ), is 6/6 = 1. For lipid oxidation, however, this is not true (e.g. tripalmitin: 2C 51H 98O 6+ 145O 2→ 102CO 2+ 98H 2O; RQ = 102/145 = 0.70) and measurement of RQ can provide useful information on substrate selection and utilisation. This will be discussed further in Chapter 11(Box 11.2).
1.3.1.1 Energy transduction
Chemical energy transduction is the process of transferring energy between different forms, and involves two main biochemical energy carrier types: (i) ATP (and also, less ubiquitously, GTP and creatine phosphate) which carries energy in the form of ‘high energy’ phosphate groups and (ii) NADH, NADPH, and FADH 2, which carry energy in the form of an electron (actually, as discussed above, a hydrogen atom with an extra electron – a hydride ion: H −). The energy status of the cell may be quantified with either of these systems – hence, (i) the ‘energy charge’ (phosphorylation potential) of the cell is an index of the degree of ATP phosphorylation (the amount of AMP that is phosphorylated to ADP and ATP: = [ATP] + [½ADP] ÷ [ATP] + [ADP] + [AMP]), and (ii) the ‘redox potential’ of the cell denotes the degree of reduction of NAD +(i.e. the NADH:NAD +ratio) and NADP +(NADPH:NADP +). Besides carrying energy ‘down’ (catabolism) or ‘up’ (anabolism) metabolic pathways ( Figure 1.12), the energy charge/phosphorylation potential and redox potential are major regulators of metabolic pathways to ensure appropriate energy provision.
1.3.1.2 Energy substrates
By utilising three, chemically diverse, fuel groups, overall metabolic flexibility and hence efficiency are achieved. Energy derived from these compounds is all based on a reduced carbon atom i.e. the C–H bond. Hence, the more C–H bonds, the more reduced the molecule and the more energy it contains, whereas oxidised or partially oxidised carbon (C–O) lacks biochemically usable energy (see Section 1.2.1.3). However, generally speaking, the more reduced the substrate, the less water soluble it is likely to be. This may be an advantage or a disadvantage, depending on the role of the substrate.
Carbohydrates are partially oxidised and hence do not contain as much energy (17 kJ g −1) as the highly reduced (–CH rich) lipids. However, carbohydrates are soluble, hence quickly mobilised and utilised, and are relatively non-toxic. Furthermore, some energy can be derived from them anaerobically during hypoxia or ischaemia. However, their water solubility means that in storage form (glycogen) they retain significant water of hydration (about three times their own weight), lowering their energy density and efficiency as energy stores (see Figure 1.10): only very limited amounts are stored (hepatic glycogen ∼ 100g only), but since they can be converted to many other substances, including lipids and intermediates of the tricarboxylic acid (TCA) cycle (also known as the Krebs cycle – see Section 1.3.1.4), they are therefore metabolically ‘flexible’ – carbohydrates are able to supply intermediary metabolites to maintain pathway integrity ( anaplerosis , again from the Greek ανα ( ana ), up, πληρω ( plero ), to fill) in contrast to lipids, oxidation of which leads to depletion of intermediary metabolites ( cataplerosis ): hence some carbohydrate is always required for metabolism to proceed efficiently, as captured in the old aphorism ‘fat burns in the fire of carbohydrate.’ This is discussed later (Box 5.3).
Fats are the most energy-dense metabolic fuels (∼37 kJ g −1): lipids are highly reduced (energetic), water-insoluble, and very energy-dense, hence their function as the principal energy store for free-living animals, and are major energy providers to most (oxidative) tissues. However, their water-insolubility makes lipids problematic and slow to mobilise, and unlike carbohydrates they cannot yield energy anaerobically – they must be oxidised, therefore cannot be used by red blood cells (erythrocytes) and renal medulla. Because they are more reduced, relatively more oxygen is required to extract energy from lipids (2.8 ATP/O 2) compared to carbohydrates (3.7 ATP/O 2) and this may be critical in high work-load oxygen-challenged tissues such as myocardium (and exercising skeletal muscle). The storage form of lipids for energy provision is triacylglycerol, which comprises three fatty acids esterified to a glycerol backbone. Being highly hydrophobic and reduced, triacylglycerols are very energy dense and a highly efficient energy store. However, triacylglycerols are relatively slow to mobilise, must be oxidised to yield energy and cannot provide energy anaerobically, and the NEFAs from which they are assembled are amphipathic (detergent- like) and hence potentially toxic in high concentrations, disrupting structural lipids especially in the central nervous system: they cannot cross the blood-brain barrier so also cannot be used by the central nervous system (more detail in Section 5.6). Furthermore, fatty acids cannot be converted into carbohydrates or proteins, limiting their metabolic flexibility.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Human Metabolism»
Представляем Вашему вниманию похожие книги на «Human Metabolism» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Human Metabolism» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.