Charles S. Cockell - Astrobiology
Здесь есть возможность читать онлайн «Charles S. Cockell - Astrobiology» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Astrobiology
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Astrobiology: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Astrobiology»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
offers an introductory text that explores the structure of living things, the formation of the elements required for life in the Universe, the biological and geological history of the Earth, and the habitability of other planets. Written by a noted expert on the topic, the book examines many of the major conceptual foundations in astrobiology, which cover a diversity of traditional fields including chemistry, biology, geosciences, physics, and astronomy.
The book explores many profound questions such as: How did life originate on Earth? How has life persisted on Earth for over three billion years? Is there life elsewhere in the Universe? What is the future of life on Earth?
is centered on investigating the past and future of life on Earth by looking beyond Earth to get the answers. Astrobiology links the diverse scientific fields needed to understand life on our own planet and, potentially, life beyond. This new second edition:
Expands on information about the nature of astrobiology and why it is useful Contains a new chapter “What is Life?” that explores the history of attempts to understand life Contains 20% more material on the astrobiology of Mars, icy moons, the structure of life, and the habitability of planets New ‘Discussion Boxes’ to stimulate debate and thought about key questions in astrobiology New review and reflection questions for each chapter to aid learning New boxes describing the careers of astrobiologists and how they got into the subject Offers revised and updated information throughout to reflect the latest advances in the field Written for students of life sciences, physics, astronomy and related disciplines, the updated edition of
is an essential introductory text that includes recent advances to this dynamic field.
Astrobiology — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Astrobiology», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
The bacterial cell wall is often surrounded by a further layer called the glycocalyx. The glycocalyx is a network of polysaccharides. A distinct, gelatinous glycocalyx is called a capsule, whereas an irregular, diffuse layer is called a slime layer. Sugars attached to the outside of cells are sometimes called extracellular polysaccharide(EPS ). The EPS acts as a protective layer of sugars that mediates interactions with surfaces. EPS allows cells to bind minerals and form biofilms. It is the substance that bonds minerals and bacteria together into macroscopic stromatolitesor domes of microorganisms. Cyanobacteriaare particularly prolific producers of EPS.
5.5.2 Archaeal Membranes
A fascinating difference in cell membranes is to be found in the archaea (Figure 5.8). This domain of microorganisms is discussed in more detail in the chapters on life in extreme environments ( Chapter 7) and phylogenetic trees ( Chapter 8). In the archaea, the lipids that make up the membranes are different from those of the bacteria and eukaryotes, one of the characteristics that put them into a separate domain of life. Instead of ester groups within the hydrophilic head of their lipids, they have ether groups, which are more resistant to a variety of chemicals and could play a role in their resistance to extreme environments. The long tails of the lipids are also different. They are called terpenoids (or isoprenoids) and have methyl side groups, in contrast to bacteria where the lipid tails are long and simple (fatty acids). These side groups might make the membranes less leaky and more resistant to extreme conditions. Even more strangely, in some archaea the bilayer is replaced by a monolayer where the tails from two lipids are fused together (Figure 5.8). This is the case in Ferroplasma , an archaeon (member of the archaea) that lives in acidic environments. It is thought that this adaptation might make the membranes more resistant to extreme conditions, preventing the membranes from falling apart.
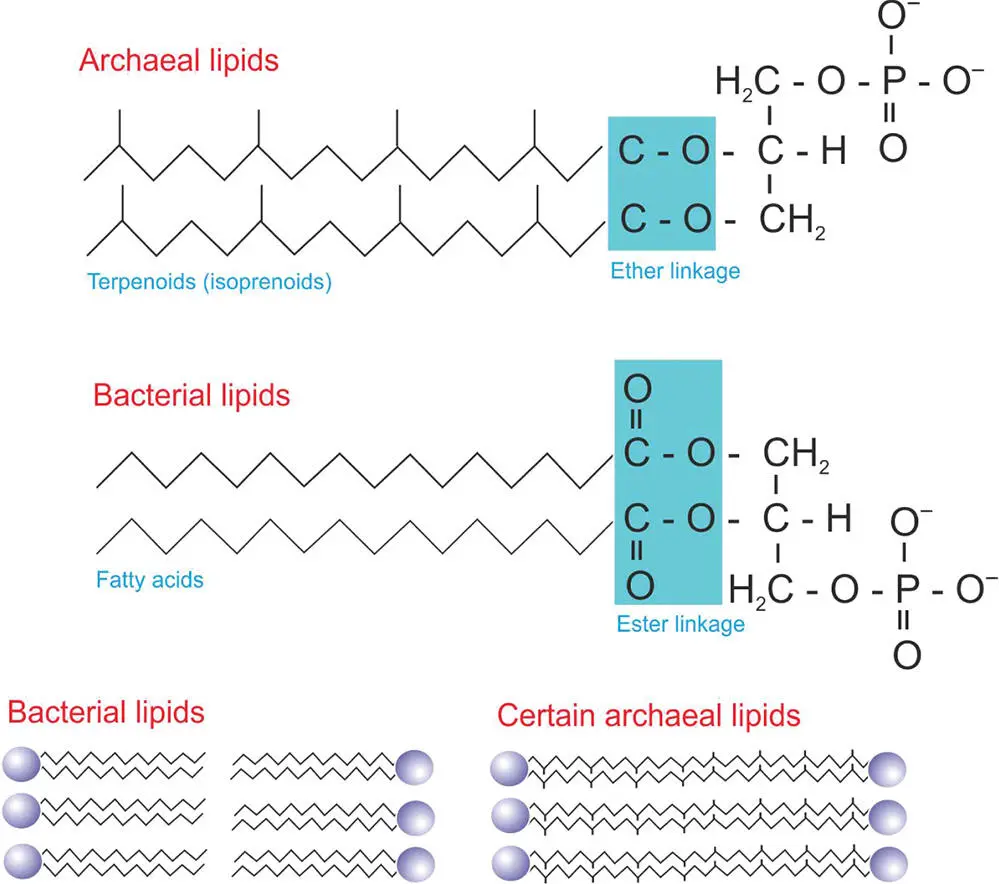
Figure 5.8 The structure of archaeal cell membrane lipids compared to bacterial lipids. The lower diagram also shows how, in some organisms, archaeal lipids can be joined in the middle.
These membrane differences between archaea and bacteria have astrobiological significance. Preserved membrane lipids can be used to tell the identity of long-dead organisms. For example, archaea are often the dominant microorganisms in salty environments. Membrane isoprenoids preserved in salts are diagnostic of archaea and can be used to infer information on the microbial communities that once lived in the salts. Different membrane lipids preserve differently. In other words, their taphonomicpotential is different. To be able to interpret which organisms lived in ancient environments, preserved in salt or rock, we need to know about cell membrane components, their ability to be preserved, and which ones might be more quickly degraded, leaving no trace.
5.6 The Information Storage System of Life
We now have a membrane containing the molecule of life. But obvious questions arise: How is the biochemistry of life controlled? What is the way in which instructions are read and directed to make molecules required for the cell to function and eventually to reproduce itself? The key information storage molecule of life is deoxyribonucleic acid , DNA. We briefly explored the structure of this molecule in the previous chapter. We learned about the four bases, guanine, adenine, cytosine, and thymine, which comprise the information code within the DNA, the bases strung together in a long sequence of different combinations. Here, we investigate how the code is turned into useful proteins.
The information storage system is crucial to the cell, since from this information, all cell instructions and coordinated biological patterns emerge. Instructions for the cell to replicate itself and interact with its environment are provided. Within complex multicellular organisms, this information coordinates cells, allowing them to communicate with each other and create differentiated cell structures, with each cell playing a dedicated role in the whole.
5.6.1 Transcription – DNA to RNA
The question arises: How do we go from the DNA information storage system to making proteins and other molecules needed by the cell for growth and reproduction?
This extraordinary transformation is accomplished in two stages. First, DNA is used to make a similar molecule called ribonucleic acid , RNA, the structure of which was described in the last chapter. This process is called transcription. RNA is similar to DNA except for three features: (i) the thymine base is replaced by uracil (U), (ii) it has a ribose sugar with a hydroxyl group (–OH) in the 2′ position, which is not present in DNA (DNA is deoxy -ribose), (iii) it generally (but not always) is single-stranded. DNA and RNA are similar in that they both contain a base, sugar, and phosphate in each structural unit.
The first step in reading the DNA is to make a copy of the sequences of the bases, or a “complementary” copy of the DNA. This copy is made out of RNA. The DNA molecule unwinds along a small part of its length (about 15 base pairs long) called the transcription bubble(Figure 5.9). At this point on the DNA, an RNA polymerase binds. RNA polymerase is a protein with five subunits and a sigma factor, a small specialized protein that recognizes particular promoterson the DNA. Promoters are sequences of DNA that correspond to the beginning of the sequences of DNA that are to be decoded. In colloquial language, the promoters tell the RNA polymerase: “Start decoding the DNA into an RNA strand here.” The RNA polymerase reads along the DNA strand, generating an RNA molecule, called messenger ribonucleic acidor mRNA. This mRNA molecule is sometimes called the mRNA transcript or primary transcript. The production of the mRNA strand occurs from the 3′ to the 5′ end of the DNA strand.
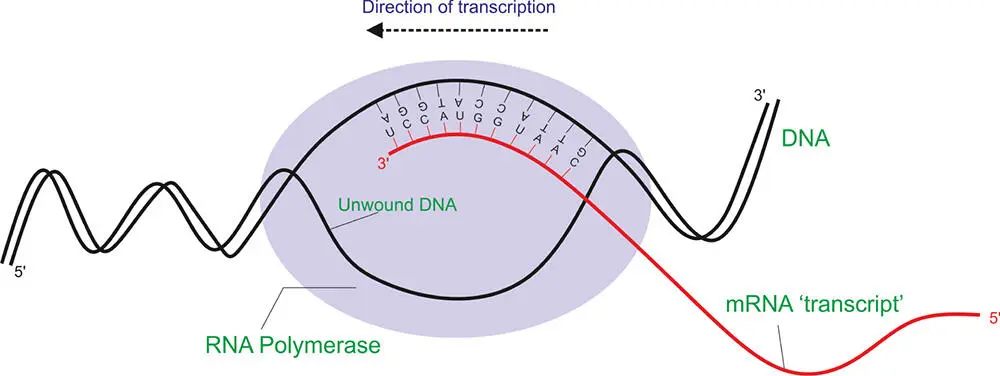
Figure 5.9 The transcription of DNA into mRNA.
In prokaryotes, this process occurs in the cytoplasm of the cell. In eukaryotes, it occurs in the cell nucleus. In eukaryotic cells, the RNA transcript must be transported out of the cell nucleus into the cytoplasm. The mRNA strand so produced can then act as the template for protein synthesis.
5.6.2 Translation – RNA to Protein
Another question now follows: How do we read the mRNA into protein? This is achieved in the next process called translation.
In translation, a new set of molecular apparatus binds to the mRNA to read the code into protein. A central part of this machinery is yet another type of RNA that is folded into a ribosome(sometimes called ribosomal ribonucleic acidor rRNA). The ribosome binds to the mRNA strand. The ribosome provides a scaffold on which other pieces of RNA called transfer ribonucleic acid( tRNA) can bind. tRNAs are the molecules that bring in the amino acids to allow for the assembly of proteins. They can be considered to be adaptor molecules that bind to the mRNA and bring amino acids into alignment to add to a growing polypeptide chain.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Astrobiology»
Представляем Вашему вниманию похожие книги на «Astrobiology» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Astrobiology» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.