Tina M. Henkin - Snyder and Champness Molecular Genetics of Bacteria
Здесь есть возможность читать онлайн «Tina M. Henkin - Snyder and Champness Molecular Genetics of Bacteria» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Snyder and Champness Molecular Genetics of Bacteria
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Snyder and Champness Molecular Genetics of Bacteria: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Snyder and Champness Molecular Genetics of Bacteria»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Although the text is centered on the most-studied bacteria,
and
, many examples are drawn from other bacteria of experimental, medical, ecological, and biotechnological importance. The book's many useful features include
Text boxes to help students make connections to relevant topics related to other organisms, including humans A summary of main points at the end of each chapter Questions for discussion and independent thought A list of suggested readings for background and further investigation in each chapter Fully illustrated with detailed diagrams and photos in full color A glossary of terms highlighted in the text While intended as an undergraduate or beginning graduate textbook, Molecular Genetics of Bacteria is an invaluable reference for anyone working in the fields of microbiology, genetics, biochemistry, bioengineering, medicine, molecular biology, and biotechnology.
"This is a marvelous textbook that is completely up-to-date and comprehensive, but not overwhelming. The clear prose and excellent figures make it ideal for use in teaching bacterial molecular genetics."—
, University of Washington
Snyder and Champness Molecular Genetics of Bacteria — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Snyder and Champness Molecular Genetics of Bacteria», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
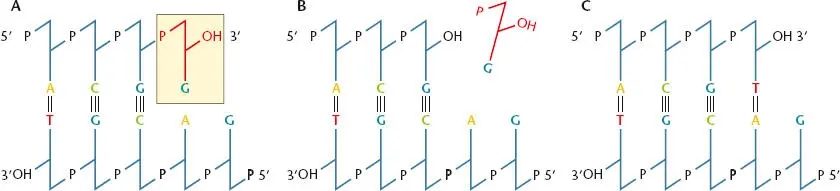
Figure 1.12 Editing function of DNA polymerase. (A)A G is mistakenly placed opposite an A while the DNA is replicating. (B and C)The DNA polymerase stops while the G is removed and replaced by a T before replication continues.
RNA Primers and Editing
The importance of the editing functions in lowering the number of mistakes during replication may explain why DNA replication is primed by RNA rather than by DNA. When the replication of a DNA chain has just initiated, the helix may be too short for distortions in its structure to be easily recognized by the editing proteins. The mistakes may then go uncorrected. However, if the first nucleotides inserted in a growing chain are ribonucleotides rather than deoxynucleotides, an RNA primer is synthesized rather than a DNA primer. The RNA primer can be removed and resynthesized as DNA by using preexisting upstream DNA as a primer. Under these conditions, a distortion in the helix can be detected by the editing functions, and mistakes are avoided.
Another important system that safeguards the fidelity of the replication process is responsible for fixing mismatches after the growing DNA strand leaves the polymerase. In E. coli and its closest relatives, this process is guided by methylation and is termed methyl-directed mismatch repair. Related mismatch repair systems are used across all three domains of life, but the use of methylation signals is not widespread. The methyl-directed mismatch repair system is discussed in chapter 10.
Impediments to DNA Replication
While the process described above and diagrammed in Figure 1.10would suffice for pristine DNA on a template that lacked any type of physical block to the progression of the DNA replication complex, in reality, the situation in the cell is rarely this tidy. DNA polymerases frequently encounter a number of different problems. Challenges to DNA polymerases include interruptions in the DNA template, bulky adducts that cannot be replicated by DNA polymerase III, and physical blocks mediated by supercoiling and proteins bound to, or acting on, the chromosome. One extreme form of impediment to a replication fork comes from nicksin either the leading-strand or lagging-strand template DNAs in which the phosphatedeoxynucleotide chain is broken in one strand of the DNA. These nicks cause the destruction of the nick-containing arm of the replication fork, resulting in a broken chromosome and collapse of the DNA replication fork. Bacteria possess a highly efficient mechanism for priming repair of this broken end by using the broken DNA itself as a primer to reinitiate DNA replication. This process was likely the original driving force for the evolution of recombination and is described in chapter 9.
Damaged DNA and DNA Polymerase III
DNA polymerase III replicates DNA with incredibly high fidelity. Much of the fidelity of the enzyme comes from the structure of the catalytic pocket, where there is a presyntheticcheck for base pairing between the template strand and the incoming nucleotide. A side effect of this small binding pocket is the inability of the polymerase to tolerate lesionsin which chemical changes have occurred in the base, the deoxyribose sugar, or even the phosphate on the DNA. There are many mechanisms for DNA replication to continue even when a cell is grown under conditions that result in highly damaged DNA. While early work suggested that the polymerization of the leading and lagging strand was so tightly coupled that a lesion on one strand would stop the entire DNA replication fork, more recent work indicates flexibility. It is now clear that although the polymerases producing the leading strand and lagging strand are physically coupled, the two complexes can be momentarily uncoupled by leaving a singlestrand DNA gap at the point of the lesion that blocked one of the DNA polymerases. Other processes can repair these gaps, and in extreme cases, where there is extensive damage in the chromosome, these gaps initiate a DNA damage response called the SOS response (see chapter 10).
Mechanisms To Deal with Impediments on Template DNA Strands
The mechanisms used for momentarily functionally uncoupling synthesis of the two strands differ depending on whether the lesion occurs on the leading-strand or lagging-strand template. The discontinuous nature of replication on the lagging-strand template affords the opportunity to circumvent lesions that halt DNA polymerase III. Typically, DNA polymerase III is recycled onto a new DNA primer when a new RNA primer is deposited ( Figure 1.10). However, a stalled lagging-strand DNA polymerase III can also be recycled by premature release when it stalls at DNA damage ( Figure 1.13A). The single-strand DNA gap left behind is repaired by another mechanism.
Under the historical model of the function of DnaG primase, primers are placed only on the lagging-strand template. However, biochemical studies indicate that in cases where the leading-strand polymerase stalls, primase can also produce an RNA primer on the leading-strand template, allowing replication to continue but leaving a gap on this strand ( Figure 1.13B). This process of lesion skipping on the leading-strand template and the ability to utilize alternative polymerases (described below) to copy over DNA damage provide complementary mechanisms to deal with damaged DNA template strands (see Gabbai et al., Suggested Reading).
While DNA polymerase III and DNA polymerase I are important for high-fidelity DNA replication, other DNA polymerases are found in E. coli that allow replication through damaged DNA in a process known as translesion synthesis. Most translesion polymerases appear to come with a trade-off in which the ability to copy damaged DNA results in a lower fidelity of DNA replication. As expected, the expression of these polymerases is induced as a response to DNA damage in the cell. In addition to controlling the amount of translesion polymerase present in the cell, access to the DNA replication fork by polymerases other than DNA polymerase III is regulated by a process called polymerase switching, a process by which one DNA polymerase replaces a polymerase already found at the 3′ OH end of a primed DNA template ( Figure 1.13C). In E. coli , DNA polymerases II, IV, and V can be recruited to temporarily step in for DNA polymerase III at damaged DNA (more details of this system are described in chapter 10). Each of these polymerases has different attributes, ranging from fairly accurate and highly processive (DNA polymerase II) to very inaccurate and not very processive (DNA polymerases IV and V). Processivity refers to how far a DNA polymerase moves on the template before falling off.
Having multiple DNA polymerases with different properties appears to be common in all living organisms. How accurate or processive a given DNA polymerase is may also depend on the nature of the damage found in the template DNA and/or the availability of various accessory proteins. The regulation of the use of these DNA polymerases is still incompletely understood, but there appear to be highly evolved processes in which the system has been fine-tuned by the process of natural selection over a long time so that the most appropriate DNA-copying mechanism is used for each environmental challenge.
Physical Blocks to Replication Forks
Proteins bound to, or otherwise acting on, the chromosome can also stop the progression of DNA polymerase III. A programmed block to DNA replication occurs in some bacteria to terminate DNA replication within one region of the chromosome (see below). However, unintended blocks can occur in other situations, such as when DNA polymerase encounters RNA polymerase carrying out transcription (see chapter 2) or when RNA polymerase is stalled at sites of damage in the chromosome. Transcription appears to be the most significant impediment to replication, but a variety of protein complexes need to be displaced ahead of the DNA replication fork.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Snyder and Champness Molecular Genetics of Bacteria»
Представляем Вашему вниманию похожие книги на «Snyder and Champness Molecular Genetics of Bacteria» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.











Обсуждение, отзывы о книге «Snyder and Champness Molecular Genetics of Bacteria» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.