Tina M. Henkin - Snyder and Champness Molecular Genetics of Bacteria
Здесь есть возможность читать онлайн «Tina M. Henkin - Snyder and Champness Molecular Genetics of Bacteria» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Snyder and Champness Molecular Genetics of Bacteria
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Snyder and Champness Molecular Genetics of Bacteria: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Snyder and Champness Molecular Genetics of Bacteria»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Although the text is centered on the most-studied bacteria,
and
, many examples are drawn from other bacteria of experimental, medical, ecological, and biotechnological importance. The book's many useful features include
Text boxes to help students make connections to relevant topics related to other organisms, including humans A summary of main points at the end of each chapter Questions for discussion and independent thought A list of suggested readings for background and further investigation in each chapter Fully illustrated with detailed diagrams and photos in full color A glossary of terms highlighted in the text While intended as an undergraduate or beginning graduate textbook, Molecular Genetics of Bacteria is an invaluable reference for anyone working in the fields of microbiology, genetics, biochemistry, bioengineering, medicine, molecular biology, and biotechnology.
"This is a marvelous textbook that is completely up-to-date and comprehensive, but not overwhelming. The clear prose and excellent figures make it ideal for use in teaching bacterial molecular genetics."—
, University of Washington
Snyder and Champness Molecular Genetics of Bacteria — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Snyder and Champness Molecular Genetics of Bacteria», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
In addition to DnaB, other helicases help DNA replication to proceed through obstacles on template DNAs. Experimentally, it was shown that two other E. coli helicases, Rep and UvrD, allow replication to proceed through a protein block on template DNA (see Guy et al., Suggested Reading). These helicases travel in the 3′-to-5′ direction, and therefore, they are likely to travel on the leading-strand template while DnaB progresses forward on the lagging-strand template (i.e., the 5′-to-3′ direction). Rep interacts directly with the DnaB helicase, probably as a normal part of the replisome. The replisome is the collection of proteins that interact with one another (through DNA or other proteins) and that are involved in carrying out DNA replication. In E. coli , UvrD may be a helicase of general use for helping out when DNA replication is blocked by proteins or to remove recombination structures from the chromosome to allow DNA replication to proceed. As explained in more detail in chapter 10, UvrD allows strand displacement during mismatch repair and plays additional roles in nucleotide excision repair in E. coli in a repair process that is coupled with transcription.
Replication of the Bacterial Chromosome and Cell Division
So far, we have discussed the details of DNA replication, but we have not discussed how bacterial DNA as a whole is replicated, nor have we discussed how the replication process is coordinated with division of the bacterial cell. To simplify the discussion, we first consider only bacteria that grow as individual cells and divide by binary fission to form two cells of equal size, even though this is far from the only type of multiplication observed among bacteria.
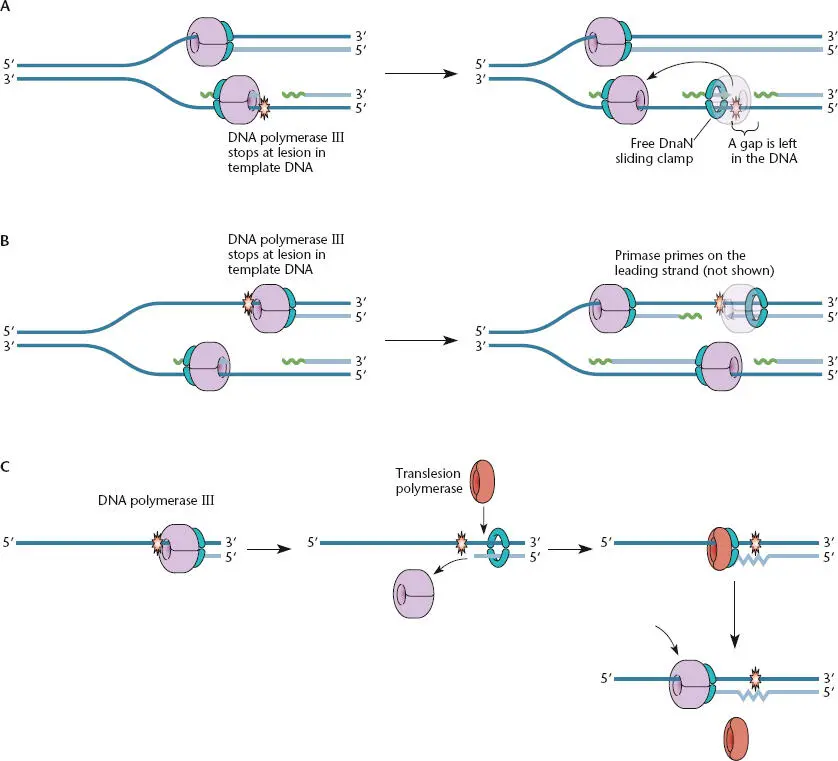
Figure 1.13 Physical blocks on template DNAs. (A)When DNA polymerase III stalls on the lagging-strand template strand, a new Okazaki fragment can be initiated and the stalled polymerase can be moved to the new RNA primer. The process leaves a gap that must be repaired by other means, probably involving RecFOR (see chapter 9). (B)When DNA polymerase III stalls on the leading strand, primase can restart DNA replication, leaving a gap that must be repaired by other means. (C)In some cases where DNA polymerase III stalls at damaged DNA, translesion polymerase can utilize the same sliding clamp to replicate through the lesion, often by error-prone DNA replication. The translesion polymerases have low processivity and fall off the template after a short distance, allowing the accurate DNA polymerase III replication to resume high-fidelity DNA replication.
The replication of the bacterial DNA occurs during the cell division cycle, which is the time during which a cell is born, grows larger, and divides into two progeny cells. Cell division is the process by which the larger cell splits into the two new cells. The division time, or generation time, is the time that elapses from the point when a cell is born until it divides. This time is usually approximately the same for all the individuals in the population under a given set of growth conditions. The original cell before cell division is called the mother cell, and the two progeny cells after division are called the daughter cells.
Structure of Bacterial Chromosomes
The DNA molecule of a bacterium that carries most of its normal genes is commonly referred to as its chromosome, by analogy to the chromosomes of higher organisms. This name distinguishes the molecule from plasmid DNA, which in some cases can be almost as large as chromosomal DNA but usually carries genes that are not always required for growth of the bacterium (see chapter 4). Most bacteria have only one chromosome; in other words, there is only one unique DNA molecule per cell that carries most of the normal genes. There are exceptions, and it is estimated that 10% of bacteria have more than one chromosome, including Vibrio cholerae , the bacterium responsible for the disease cholera. Even in bacteria that contain multiple chromosomes, the second chromosome shows more characteristics of a plasmid than of a chromosome, particularly in how it initiates replication. There appear to be special molecular systems for managing multiple chromosomes (see Fournes et al., Suggested Reading).
As discussed below, when bacteria, such as E. coli , are reproducing very rapidly, new rounds of replication initiate before others are completed, temporally increasing the DNA content of the cells until cellular division returns the number of copies to one unit chromosome per cell. It is important to note, however, that these individual chromosomal DNAs are not unique since they are directly derived from each other by replication.
The structure of bacterial DNA differs significantly from that of the chromosomes of higher organisms. One difference is that the DNA in the chromosomes of most bacteria is circular in the sense that the ends are joined to each other (for exceptions, see Box 4.1). In contrast, eukaryotic chromosomes are usually linear with free ends. As discussed in chapter 4, the circularity of bacterial chromosomal DNA allows it to replicate in its entirety without using telomeres, as eukaryotic chromosomes do, or terminally redundant ends, as some bacteriophages do (see chapter 7). Even in cases where bacterial chromosomes are linear, they do not use the same mechanism, involving telomerases to replicate their ends, that is used by eukaryotic chromosomes. Another difference between the DNA of bacteria and eukaryotes is that the DNA in eukaryotes is wrapped around proteins called histones to form nucleosomes. Bacteria have the proteins HU, HN-S, Fis, and IHF, around which DNA is often wrapped, and archaea do have rudimentary histones related to those of eukaryotes. However, in general, DNA is much less structured in bacteria than in eukaryotes.
Replication of the Bacterial Chromosome
Replication of a circular bacterial chromosome initiates at a unique origin of chromosomal replication, or oriC , and proceeds in both directions around the circle. On the E. coli chromosome, oriC is located at 84.3 min. As mentioned above, the place in DNA at which replication occurs is known as the replication fork. Two replication forks start at oriC and proceed around the circle until they meet and terminatechromosomal replication. The DNA polymerases responsible for replicating the leading and lagging strands associate as a single holoenzyme. However, there is no association between the DNA polymerases at the two DNA replication forks to help drive the separation of chromosomes, and therefore, other force-generating mechanisms must be at play. As discussed in “Termination of Chromosome Replication” below, some bacteria actively terminate replication at a unique site in the DNA; however, these systems are not widespread, and most bacteria terminate replication using an unknown mechanism or simply terminate DNA replication where the two replication forks meet. Each time the two replication forks proceed around the circle and meet, a round of replicationhas been completed, and two new DNAs, called the daughter DNAs, are generated.
Initiation of Chromosome Replication
Much has been learned about the molecular events occurring during the initiation of replication. Some of this information has a bearing on how the initiation of chromosome replication is regulated and serves as a model for the interaction of proteins and DNA.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Snyder and Champness Molecular Genetics of Bacteria»
Представляем Вашему вниманию похожие книги на «Snyder and Champness Molecular Genetics of Bacteria» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.











Обсуждение, отзывы о книге «Snyder and Champness Molecular Genetics of Bacteria» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.