Jane Flint - Principles of Virology
Здесь есть возможность читать онлайн «Jane Flint - Principles of Virology» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.
- Название:Principles of Virology
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Principles of Virology: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Principles of Virology»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Volume I: Molecular Biology
Volume II: Pathogenesis and Control
Principles of Virology, Fifth Edition
Principles of Virology — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Principles of Virology», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Phylogenetic trees can also be constructed by grouping sampled viruses by host of isolation. Such an arrangement sometimes makes it possible to identify the animal source of a human virus. Circular forms, such as a radial format tree, are often displayed when the root is unknown.
Trees relating nucleic acid sequences depict the relationships as if sampled and intermediary sequences were on a trajectory to the present. This deduction is an oversimplification, because any intermediate that was lost during evolution will not be represented in the tree. In addition, any recombination or gene exchange by coinfection with similar viral genomes will scramble ordered lineages.
A fair question is whether we can predict the future trajectory or branches of the tree. We can never answer this question for two reasons: any given sample may not represent the diversity of any given virus population in an ecosystem, and we cannot predict the selective pressures that will be imposed.
Hall BG. 2011. Phylogenetic Trees Made Easy: A How- to Manual, 4th ed. Sinauer Associates, Sunderland, MA.
ViralZone. Phylogenetics of animal pathogens: basic principles and applications (a tutorial). http://viralzone.expasy.org/e_learning/phylogenetics/content.html
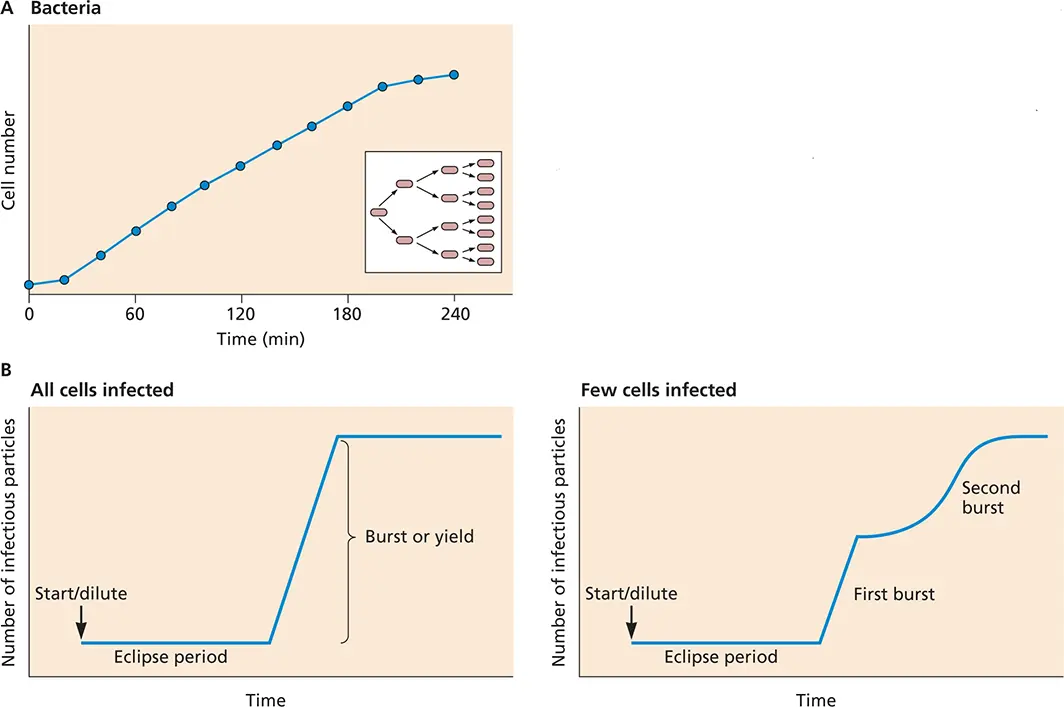
Figure 2.18 Comparison of bacterial and viral reproduction. (A)Growth curve for a bacterium. The number of bacteria is plotted as a function of time. One bacterium is added to the culture at time zero; after a brief lag, the bacterium begins to divide. The number of bacteria doubles every 20 min until nutrients in the medium are depleted and the growth rate decreases. The inset illustrates the propagation of bacteria by binary fission. (B)One- and two-step growth curves of bacteriophages. Growth of a bacteriophage in E. coli under conditions when all cells are infected (left) and when only a few cells are infected (right).
The results of these experiments defined two new terms in virology: the eclipse period, the phase in which infectivity is lost when virions are disassembled after penetrating cells, and the latent period, the time it takes to replicate and assemble new virus particles before lysis, ∼20 to 25 min for some E. coli bacteriophages.
Synchronous infection, the key to the one-step growth cycle, is usually accomplished by infecting cells with a sufficient number of virus particles to ensure that most of the cells are infected rapidly. Exactly how many virus particles must be added is described by the multiplicity of infection( Box 2.11).
One-Step Growth Analysis: a Valuable Tool for Studying Animal Viruses
One-step growth analysis soon became adapted for studying the reproduction of animal viruses. The experiment begins with removal of the medium from the cell monolayer and addition of virus in a small volume to promote rapid adsorption. After ∼1 h, unadsorbed inoculum containing virus particles is removed, the cells are washed, and fresh medium is added. At different times after infection, samples of the cell culture supernatant are collected and the virus titer is determined. The kinetics of intracellular virus production can be monitored by removing the medium containing extracellular particles, scraping the cells into fresh medium, and lysing them. A cell extract is prepared after removal of cellular debris by centrifugation, and the virus titer in the extract is measured.
The results of a one-step growth experiment establish a number of important features about viral reproduction. In the example shown in Fig. 2.19A, the first 11 h after infection constitute the eclipse period, during which the viral nucleic acid is uncoated from its protective shell and no infectious virus can be detected inside cells. The small number of infectious particles detected during this period probably represents adsorbed virus that was not uncoated. Beginning at 12 h after adsorption, the quantity of intracellular infectious virus particles begins to increase, marking the onset of the synthetic phase, during which assembly begins. During the latent period, no extracellular virus can be detected. At 18 h after adsorption, virions are released from cells into the extracellular medium. Ultimately, virus numbers plateau as the cells become metabolically and structurally incapable of supporting additional reproduction.
BOX 2.11
DISCUSSION
Multiplicity of infection (MOI)
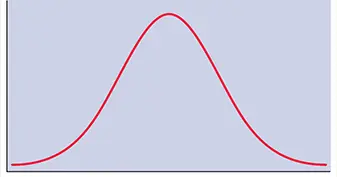
Infection depends on the random collision of cells and virus particles. When susceptible cells are mixed with a suspension of virus particles, some cells are uninfected and other cells receive one, two, three, etc., particles. The distribution of virus particles per cell is best described by the Poisson distribution:
P(k) = e –m mk/k!
In this equation, P ( k ) is the fraction of cells infected by k virus particles. The multiplicity of infection, m , is calculated from the proportion of uninfected cells, P (0), which can be determined experimentally. If k is made 0 in the above equation, then
P (0) = e –mand m = −1 n P (0)
The fraction of cells receiving 0, 1, and >1 virus particle in a culture of 10 6cells infected with an MOI of 10 can be determined as follows.
The fraction of cells that receive 0 particles is
P (0) = e–10 = 4.5 × 10 –5
and in a culture of 10 6cells, this equals 45 uninfected cells.
The fraction of cells that receive 1 particle is
P (1) = 10 × 4.5 × 10 –5= 4.5 × 10 –4
and in a culture of 10 6cells, 450 cells receive 1 particle.
The fraction of cells that receive >1 particle is
P (>1) = 1 − e –m( m + 1 ) = 0.9995
and in a culture of 10 6cells, 999,500 cells receive >1 particle. [The value in this equation is obtained by subtracting from 1 (the sum of all probabilities for any value of k ) the probabilities P (0) and P (1).]
The fraction of cells receiving 0, 1, and >1 virus particle in a culture of 10 6cells infected with an MOI of 0.001 is
1 P(0) = 99.99%P(1) = 0.0999% (for 106 cells, 104 are infected)P(>1) = 10–6The MOI required to infect 99% of the cells in a cell culture dish isP(0) = 1% = 0.01m = −1n (0.01) = 4.6 PFU per cell.
The yield of infectious virus per cell can be calculated from the data collected during a one-step growth experiment. This value varies widely among different viruses and with different virus-host cell combinations. For many viruses, increasing the multiplicity of infection above a certain point does not increase the yield: cells have a finite capacity to produce new virus particles. In fact, infecting at a very high multiplicity of infection can cause premature cell lysis and decrease virus yields.
The kinetics of the one-step infectious cycle can vary dramatically among viruses. For example, enveloped viruses that mature by budding from the plasma membrane, as discussed in Chapter 13, generally become infectious only as they leave the cell, and therefore, little intracellular infectious virus can be detected ( Fig. 2.19B). The curve shown in Fig. 2.19Aillustrates the pattern observed for a DNA virus with the long latent and synthetic phases typical of many DNA viruses, some retroviruses, and reovirus. For small RNA viruses, the entire growth curve is complete within 6 to 8 h, and the latent and synthetic phases are correspondingly shorter.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Principles of Virology»
Представляем Вашему вниманию похожие книги на «Principles of Virology» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Principles of Virology» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.