George Acquaah - Principles of Plant Genetics and Breeding
Здесь есть возможность читать онлайн «George Acquaah - Principles of Plant Genetics and Breeding» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Principles of Plant Genetics and Breeding
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Principles of Plant Genetics and Breeding: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Principles of Plant Genetics and Breeding»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Organizes topics to reflect the stages of an actual breeding project Incorporates the most recent technologies in the field, such as CRSPR genome edition and grafting on GM stock Includes numerous illustrations and end-of-chapter self-assessment questions, key references, suggested readings, and links to relevant websites Features a companion website containing additional artwork and instructor resources
offers researchers and professionals an invaluable resource and remains the ideal textbook for advanced undergraduates and graduates in plant science, particularly those studying plant breeding, biotechnology, and genetics.
Principles of Plant Genetics and Breeding — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Principles of Plant Genetics and Breeding», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Whereas outcrossing species have more heterozygous loci and carry more unfitness load, there are cases in which the heterozygote is fitter than either homozygote. Called overdominance, this phenomenon is exploited in hybrid breeding (see Chapter 18).
3.9 Inbreeding and its implications in plant breeding
The point has already been made that the methods used by plant breeders depend on the natural means of reproduction of the species. This is because each method of reproduction has certain genetic consequences. In Figure 3.3a, there is no inbreeding because there is no common ancestral pathway to the individual, A (i.e. all parents are different). However, in Figure 3.3b inbreeding exists because B and C have common parents (D and E), that is, they are full sibs. To calculate the amount of inbreeding, the standard pedigree is converted to an arrow diagram ( Figure 3.3c). Each individual contributes ½ of its genotype to its offspring. The coefficient of relationship (R) is calculated by summing up all the pathways between two individuals through a common ancestor as: R BC= Σ(½) s, where s is the number of steps (arrows) from B to the common ancestor and back to C. For example, B and C probably inherited (½)(½) = ¼ of their genes in common through ancestor D. Similarly, B and C probably inherited ¼ of their genes in common through ancestor E. The coefficient of relationship between B and C, as a result of common ancestry, is hence R BC= ¼ + ¼ = ½ = 50%. Other more complex pedigrees are shown in Figure 3.4.
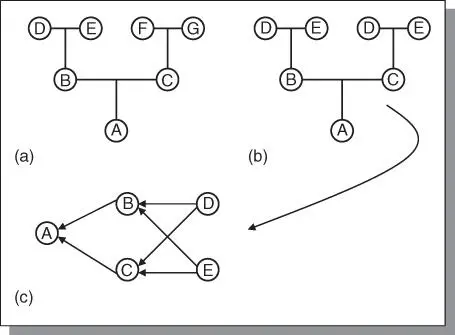
Figure 3.3Pedigree diagrams can be drawn in the standard form (a or b) or converted to into an arrow diagram (c).
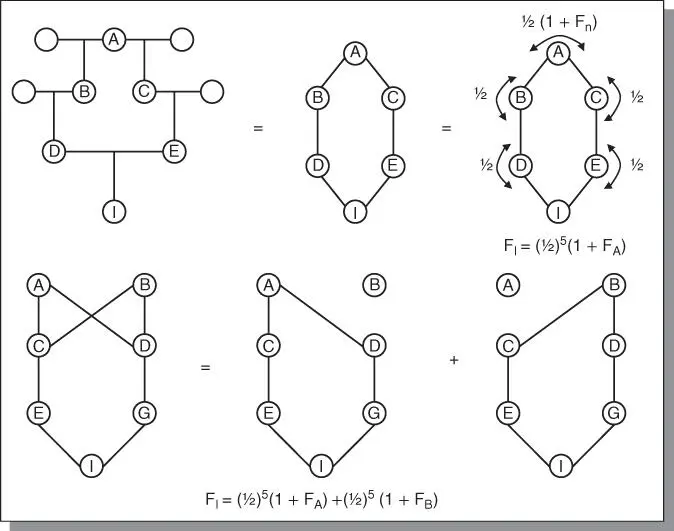
Figure 3.4The inbreeding coefficient may be calculated by counting the number of arrows which connect the individual through one parent back to the common ancestor and back again to the other parent and applying the formula in the figure.
As previously indicated, prolonged selfing is the most extreme form of inbreeding. With each selfing, the percent heterozygosity decreases by 50%, whereas the percent homozygosity increases by 50% from the previous generation. The approach to homozygosity depends on the intensity of inbreeding as illustrated in Figure 3.5. The more distant the relationship between parents, the slower is the approach to homozygosity. The coefficient of inbreeding (F), previously discussed, measures the probability of identity of alleles by descent. This can be measured at both the individual level as well as the population level. At the individual level, F measures the probability that any two alleles at any locus are identical by descent (i.e. they are both products of a gene present in a common ancestor) At the population level, F measures the percentage of all loci which were heterozygous in the base population but have now probably become homozygous due to the effects of inbreeding. There are several methods used for calculating F. The coefficient of inbreeding (Fx) of an individual may be obtained by counting the number of arrows (n) that connect the individual through one parent back to the common ancestor and back again to the other parent, and using the mathematical expression:
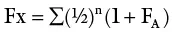
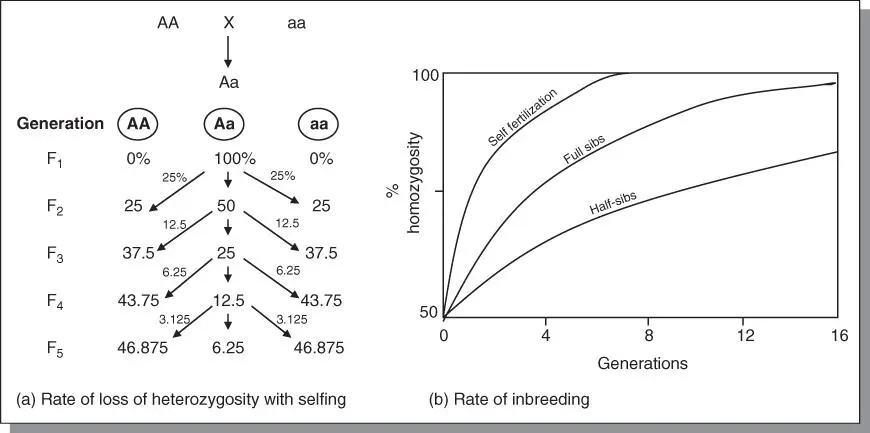
Figure 3.5Increase in percentage of homozygosity under various systems of inbreeding. (a) Selfing reduces heterozygosity by 50% of what existed at the previous generation. (b) The approach to homozygosity is most rapid under self‐fertilization.
3.9.1 Consequences
The genetic consequences of inbreeding were alluded to in Section 3.8. The tendency toward homozygosity with inbreeding provides an opportunity for recessive alleles to be homozygous and hence expressed. Whereas inbreeding generally has little or no adverse effect in inbred species, crossbred species suffer adverse consequences when the recessive alleles are less favorable than the dominant alleles. Called inbreeding depression, it is manifested as a reduction in performance, because of the expression of less fit or deleterious alleles. The severity of inbreeding depression varies among species, being extreme in species such as alfalfa in which inbreeding produces homozygous plants that fail to survive. Further, the effect of inbreeding is most significant in the first five to eight generations, and negligible after the eighth generation in many cases.
3.9.2 Applications
Inbreeding is desirable in some breeding programs. Inbred cultivars of self‐pollinated species retain their genotype through years of production. In cross‐pollinated species, inbred lines are deliberately developed for use as parents in hybrid seed production. Similarly, partially inbred lines are used as parents in the breeding of synthetic cultivars and vegetatively propagated species by reducing the genetic load. Another advantage of inbreeding is that it increases the genetic diversity among individuals in a population, thereby facilitating the selection process in a breeding program.
3.9.3 Mating systems that promote inbreeding
Mating is a way by which plant breeders impact the gene frequencies in a population. Four mating systemsare commonly used to effect inbreeding: self‐fertilization, full‐sib mating, half‐sib mating, and backcrossing. Self‐fertilization is the union of male and female gametes; full‐sib mating involves the crossing of pairs of plants from a population. In half‐sib mating, the pollen source is random from the population, but the female plants are identifiable. In a backcross, the F 1is repeatedly crossed to one of the parents. Self‐fertilization and backcrossing are the most extreme forms of inbreeding attaining a coefficient of inbreeding (F) of 15/16 after four generations of mating. Autopolyploids have multiple alleles and hence can accumulate more deleterious alleles that remain masked. Inbreeding depression is usually more severe in autopolyploids than diploid species. However, the progression to homozygosity is much slower in autopolyploids than in diploids.
3.10 Concept of population improvement
The general goal of improving open or cross‐pollinated species is to change the gene frequencies in the population toward fixation of favorable alleles while maintaining a high degree of heterozygosity. Unlike self‐pollinated species in which individuals are the focus and homozygosity and homogeneity are desired outcomes of breeding, population improvement focuses on the whole group, not individual plants. Consequently, open‐pollinated populations are not homogeneous.
3.11 Types
The population can be changed by one of two general strategies (i.e. there are two basic types of open pollinated populations in plant breeding) – by population improvementand by development of synthetic cultivars. To develop cultivars by population improvement entails changing the population en masse by implementing a specific selection tactic. A cultivar developed this way is sustainable in a sense, maintaining its identity indefinitely through random mating within itself in isolation. The terminology “synthetic” is used to denote an open pollinated cultivar developed from combining inbred or clonal parental lines. However, the cultivar is not sustainable and must be reconstituted from parental stock. Other usage of the term occurs in the literature.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Principles of Plant Genetics and Breeding»
Представляем Вашему вниманию похожие книги на «Principles of Plant Genetics and Breeding» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Principles of Plant Genetics and Breeding» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.