Michael Begon - Ecology
Здесь есть возможность читать онлайн «Michael Begon - Ecology» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Ecology
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Ecology: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Ecology»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
– now in full colour – offers students and practitioners a review of the ecological sciences.
The previous editions of this book earned the authors the prestigious ‘Exceptional Life-time Achievement Award’ of the British Ecological Society – the aim for the fifth edition is not only to maintain standards but indeed to enhance its coverage of Ecology.
In the first edition, 34 years ago, it seemed acceptable for ecologists to hold a comfortable, objective, not to say aloof position, from which the ecological communities around us were simply material for which we sought a scientific understanding. Now, we must accept the immediacy of the many environmental problems that threaten us and the responsibility of ecologists to play their full part in addressing these problems. This fifth edition addresses this challenge, with several chapters devoted entirely to applied topics, and examples of how ecological principles have been applied to problems facing us highlighted throughout the remaining nineteen chapters.
Nonetheless, the authors remain wedded to the belief that environmental action can only ever be as sound as the ecological principles on which it is based. Hence, while trying harder than ever to help improve preparedness for addressing the environmental problems of the years ahead, the book remains, in its essence, an exposition of the
of ecology. This new edition incorporates the results from more than a thousand recent studies into a fully up-to-date text.
Written for students of ecology, researchers and practitioners, the fifth edition of
is anessential reference to all aspects of ecology and addresses environmental problems of the future.
Ecology — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Ecology», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Even when generations overlap, if individuals can be marked early in their life so that they can be recognised subsequently, it is feasible to follow the fate of each year’s cohort separately. It may then be possible to merge the cohorts from the different years of a study to derive a single, ‘typical’ cohort life table. An example is shown in Table 4.2of females from a population of the yellow‐bellied marmot, Marmota flaviventris . The population was live‐trapped and marmots marked individually from 1962 through to 1993 in the East River Valley of Colorado, USA and it was this that allowed each individual to be assigned, whenever it was caught, to its own cohort.
Table 4.2 A cohort life table for female yellow‐bellied marmots, Marmota flaviventris in Colorado, USA.The columns are explained in the text. Source : After Schwartz et al . (1998).
| Age class (years) x | Number alive at the start of each age class ax | Proportion of original cohort surviving to the start of each age class lx | dx | qx | log ax | log lx | kx | Number of female young produced by each age class Fx | Number of female young produced per surviving individual in each age class mx | Number of female young produced per original individual in each age class lxmx |
|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 773 | 1.000 | 0.457 | 0.457 | 2.888 | 0.00 | 0.26 | 0 | 0.000 | 0.000 |
| 1 | 420 | 0.543 | 0.274 | 0.505 | 2.623 | −0.26 | 0.31 | 0 | 0.000 | 0.000 |
| 2 | 208 | 0.269 | 0.089 | 0.332 | 2.318 | −0.57 | 0.18 | 95 | 0.457 | 0.123 |
| 3 | 139 | 0.180 | 0.043 | 0.237 | 2.143 | −0.75 | 0.12 | 102 | 0.734 | 0.132 |
| 4 | 106 | 0.137 | 0.050 | 0.368 | 2.025 | −0.86 | 0.20 | 106 | 1.000 | 0.137 |
| 5 | 67 | 0.087 | 0.030 | 0.343 | 1.826 | −1.06 | 0.18 | 75 | 1.122 | 0.098 |
| 6 | 44 | 0.057 | 0.017 | 0.295 | 1.643 | −1.24 | 0.15 | 45 | 1.020 | 0.058 |
| 7 | 31 | 0.040 | 0.012 | 0.290 | 1.491 | −1.40 | 0.15 | 34 | 1.093 | 0.044 |
| 8 | 22 | 0.029 | 0.013 | 0.455 | 1.342 | −1.55 | 0.26 | 37 | 1.680 | 0.049 |
| 9 | 12 | 0.016 | 0.006 | 0.417 | 1.079 | −1.81 | 0.23 | 16 | 1.336 | 0.021 |
| 10 | 7 | 0.009 | 0.005 | 0.571 | 0.845 | −2.04 | 0.37 | 9 | 1.286 | 0.012 |
| 11 | 3 | 0.004 | 0.001 | 0.333 | 0,477 | −2.41 | 0.18 | 0 | 0.000 | 0.000 |
| 12 | 2 | 0.003 | 0.000 | 0.000 | 0.301 | −2.59 | 0.00 | 0 | 0.000 | 0.000 |
| 13 | 2 | 0.003 | 0.000 | 0.000 | 0.301 | −2.59 | 0.00 | 0 | 0.000 | 0.000 |
| 14 | 2 | 0.003 | 0.001 | 0.500 | 0.301 | −2.59 | 0.30 | 0 | 0.000 | 0.000 |
| 15 | 1 | 0.001 | 0.000 | −2.89 | 0 | 0.000 | 0.000 | |||
| Total | 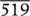 |
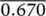 |
the columns of a life table
The first column in each life table is a list of the stages or age classes of the organism’s life. For Gilia , these are simply the stages ‘seed’, ‘emerged plants’, and ‘flowering plants’. For the marmots, they are years. The second column is then the raw data from each study, collected in the field. It reports the number of individuals surviving to the beginning of each stage or age class (see Figure 4.9). We refer to these numbers as a x, where the x in the subscript refers to the stage or age class concerned: a 0means the numbers in the initial age class, and so on.
Ecologists are typically interested not just in examining populations in isolation but in comparing the dynamics of two or more perhaps rather different populations. This was precisely the case for the Gilia populations in Table 4.1. Hence, it is necessary to standardise the raw data so that comparisons can be made. This is done in the third column of the table, which is said to contain l xvalues, where l xis defined as the proportion of the original cohort surviving to the start of age class. The first value in this column, l 0(spoken: L‐zero), is therefore the proportion surviving to the beginning of this original age class. Obviously, in Tables 4.1and 4.2, and in every life table, l 0is 1.00 (the whole cohort is there at the start). Thereafter, in the marmots for example, there were 773 females observed in this youngest age class. The l xvalues for subsequent age classes are therefore expressed as proportions of this number. Only 420 individuals survived to reach their second year (age class 1: between one and two years of age). Thus, in Table 4.2, the second value in the third column, l 1, is the proportion 420/773 = 0.543 (that is, only 0.543 or 54.3% of the original cohort survived this first step). In the next row, l 2= 208/773 = 0.269, and so on. For Gilia ( Table 4.1), l 1= 254/746 = 0.340 for the inland subspecies and 204/754 = 0.271 for the coastal subspecies. That is, 34% and 27.1% survived the first step to become established plants in the two cases: a slightly higher survival rate at this inland site for the inland than for the coastal subspecies.
In the next column, to consider mortality more explicitly, the proportion of the original cohort dying during each stage ( d x) is computed, being simply the difference between successive values of l x; for example, for the marmots, d 3= l 3– l 4= 0.180 − 0.137 = 0.043. Next, the stage‐specific mortality rate, q x, is computed. This considers d xas a fraction of l x. Hence, q 3for example is 0.24 (= 0.043/0.180 or d 3/ l 3). Values of q xmay also be thought of as the average ‘chances’ or probabilities of an individual dying during an interval. q xis therefore equivalent to (1 − p x) where p refers to the probability of survival.
The advantage of the d xvalues is that they can be summed: thus, the proportion of a cohort of marmots dying in the first four years was d 0+ d 1+ d 2+ d 3(= 0.86). The disadvantage is that the individual values give no real idea of the intensity or importance of mortality during a particular stage. This is because the d xvalues are larger the more individuals there are, and hence the more there are available to die. The q xvalues, on the other hand, are an excellent measure of the intensity of mortality. For instance, in the present example it is clear from the q xcolumn that the mortality rate declined after the first two years of life but then rose again to a peak around years 9 and 10; this is not clear from the d xcolumn. The q xvalues, however, have the disadvantage that, for example, summing the values over the first four years gives no idea of the mortality rate over that period as a whole.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Ecology»
Представляем Вашему вниманию похожие книги на «Ecology» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Ecology» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.