An Introduction to Molecular Biotechnology
Здесь есть возможность читать онлайн «An Introduction to Molecular Biotechnology» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:An Introduction to Molecular Biotechnology
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
An Introduction to Molecular Biotechnology: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «An Introduction to Molecular Biotechnology»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
An Introduction to Molecular Biotechnology — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «An Introduction to Molecular Biotechnology», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
In meiosisthe sets of chromosomes are divided in half ( reduction division), therefore resulting in the haploid genome. Meiosis is required for sexual reproduction in diploid organisms. If the gametes were diploid, every new zygote formed would contain double the number of chromosomes. This dilemma can only be solved through haploid gametes. Meiosis also serves to mix the paternal genes and increases genetic variability, which is an important prerequisite for natural selection.
The most important difference between meiosis and mitosis lies in the pairing of homologous chromosomes and the consequent reduction of the chromosome set. Meiosisis divided into two divisions: the first and second reduction division (or meiosis Iand meiosis II). In prophaseof the first reduction division, there are five stages: leptotene, zygotene, pachytene, diplotene, and diakinesis. After doubling of the chromosomes, two sister chromatids become apparent in the leptotene. In zygotenethe pairing of the homologous maternal and paternal chromosomes begins ( synapsis). The chromosomes paired in each case correspond to each other at the sequence level, enabling crossing‐overand recombinationto take place. These processes enhance the exchange of genetic information from both parents. In pachytenethe pairing of homologous chromosomes is completed. In the following diplotenethe chromosome pairs separate from one another but cling together at the sites on which crossing‐over took place (so‐called chiasmata). In this phase, the chromosomes unwind and are transcriptionally active. In diakinesisthe transcription ends, and the chromosomes condense again.
In the following metaphase I ,the nuclear membrane and nucleolus break down, and the spindle apparatus is formed. The chromosome pairs assemble themselves on the equator so that the centromeres are orientated to the spindle poles. However, the kinetochore microtubules do not attach to the centromeres of individual chromatids (as in mitosis), but to a shared centromere of each chromatid pair. In meiotic anaphase I, the chromosome pairs are pulled apart over the shortening kinetochore microtubules to the cell poles. The recombinant chromosome areas separate at the chiasmata. The sister chromatids remain joined together via the centromere.
After a short interphase, the second reduction division occurs. This division contains the mechanics of mitosis. The chromosomes are ordered again as in metaphase: metaphase II. The chromatids are pulled apart by the kinetochore microtubules to the cell poles. This process is completed with anaphase I I. After the telophase II and cytokinesis, haploid cells remain (so‐called meiosporesor meiogametes), each with a haploid chromosome set available. During the formation of haploid gametes, the chromosomes from the parents are sorted randomly, which again increases genetic variability.
4.1.4 Replication
With every cell division(mitosis; Figure 4.7), the entire genome of the cell is duplicated. This means that two identical chromatidsresult from every chromosome. These chromatids are identical daughter chromosomes that will be distributed to the daughter cells following cell division. The duplication of the DNA, which is referred to as DNA replication, occurs in a semiconservative manner.
During semiconservative replication, the DNA double strand is locally separated into single strands, and a replication forkis formed. The single strands serve as a matrix for the synthesis of each complementary new strand. The DNA replication is a complex process, in which many proteins and enzymes are involved ( Figure 4.8). To open the double strands, a helicaseis needed. The leading strand, orientated in the 3′ → 5′ direction, can be copied directly by the DNA polymerase, as synthesis occurs in a 5′ → 3′ direction. The opposing strand, termed lagging strand, cannot be copied in the same way as it is orientated in a 5′ → 3′ direction. As soon as the DNA is present as single strands, specific proteins bind ( single‐strand binding proteins) and prevent the reformation of the double helix. DNA primaseplaces short RNA primers, which are complementary to the DNA sequence, at regular intervals on the lagging strand. These RNA primers can be lengthened by DNA polymerase until the next RNA primer is reached (referred to as Okazaki fragments). The RNA is then removed and replaced by deoxynucleoside triphosphates (dNTPs). The Okazaki fragments are linked through the DNA ligase. A sliding clampkeeps the DNA polymerase firmly on the DNA when it is moving. An ATP‐depending clamp loaderprotein complex supports this process. Enzymes involved in replication vary between prokaryotes and eukaryotes. However, the general composition of the multienzyme complex is similar. During the unwinding of the double helix, DNA topoisomerases cut the DNA at regular intervals to prevent the rotation of the double helix. DNA topoisomerase Icatalyzes single‐strand breaking, while DNA topoisomerase IIcan cut both DNA strands at the same time.
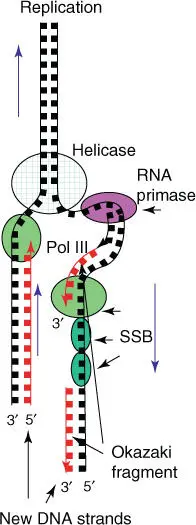
Figure 4.8 Schematic summary of DNA replication. SSB, single‐strand binding proteins; Pol III, DNA polymerase III.
Replication begins at specific DNA sequences termed origins. Here the replication bubbleopens, and replication occurs in parallel on both the right and left replication forks ( Figure 4.9). Whereas in circular bacterial genomes, only one origin of replicationis present; in eukaryotic chromosomesa replication start site is positioned on the linear chromosomes every 1000 bp. In this way, even long chromosomes can be replicated in a short time. In eukaryotic cells, four phases are distinguished in a cell cycle: the S phase(DNA synthesis) lasts around eight hours in mammalian cells. The replicated chromatids stay together until the M phase(when mitosis starts). S and M phases are separated by G1 and G2 phases(G for gap). More details of the cell cycle and its measurement can be found in Chapter 18.
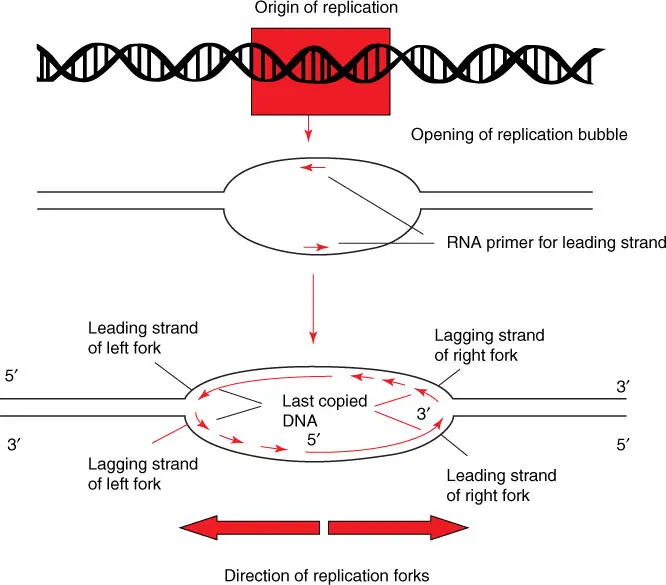
Figure 4.9 Asymmetric composition of replication bubbles. DNA is unwound at the origin of replication, and a replication bubble with a right and left replication fork is formed. Replication proceeds in parallel within the replication bubble. DNA primase introduces a complementary RNA primer on each leading strand, so that DNA polymerase III can carry out replication. The individual lagging stands are synthesized as shown in Figure 4.8.
DNA polymerasescopy the original nucleotide sequence flawlessly (the error rate during synthesis is one incorrect nucleotide per 10 000 nucleotides). However, special repair enzymesplay a large role. Incorrectly paired nucleotides are removed by specific exonucleasesand then replaced through DNA polymerase; finally, the phosphoester bond is covalently linked through DNA ligase. Together, these mechanisms reduce mutation rates to less than one in 10 billion nucleotides. Thus, DNA synthesis displays a very high fidelity.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «An Introduction to Molecular Biotechnology»
Представляем Вашему вниманию похожие книги на «An Introduction to Molecular Biotechnology» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.


![Andrew Radford - Linguistics An Introduction [Second Edition]](/books/397851/andrew-radford-linguistics-an-introduction-second-thumb.webp)









Обсуждение, отзывы о книге «An Introduction to Molecular Biotechnology» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.