An Introduction to Molecular Biotechnology
Здесь есть возможность читать онлайн «An Introduction to Molecular Biotechnology» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:An Introduction to Molecular Biotechnology
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
An Introduction to Molecular Biotechnology: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «An Introduction to Molecular Biotechnology»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
An Introduction to Molecular Biotechnology — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «An Introduction to Molecular Biotechnology», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
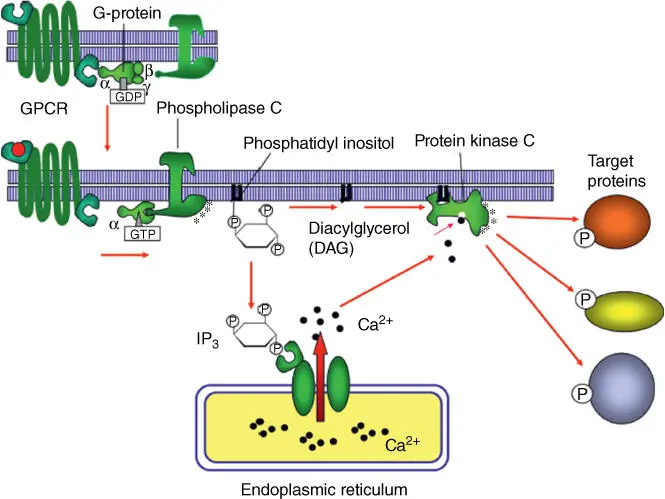
Figure 3.10 Role of phospholipase C‐ β in the production of second messengers IP 3and DAG.
Source: Alberts et al. (2015). Adapted with permission of Garland Science.
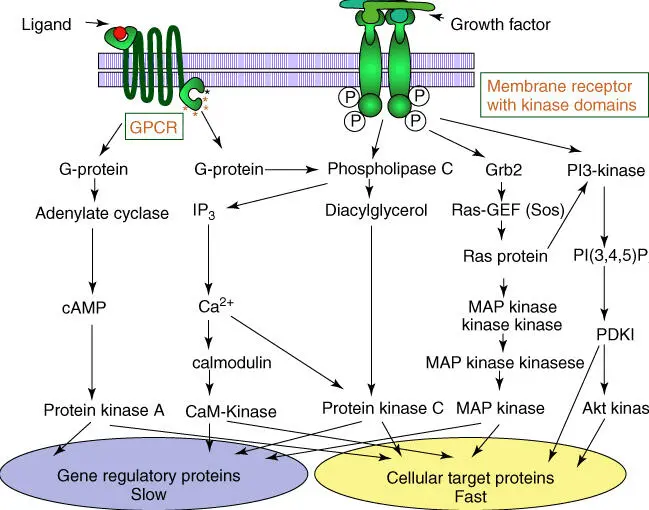
Figure 3.11 Signal transduction after activation of G‐protein and enzyme‐linked receptors. GPCR, G‐protein‐coupled receptor; GEF, guanine exchange factor.
Source: Alberts et al. (2015). Adapted with permission of Garland Science.
Table 3.5 Signal proteins that act via receptor tyrosine kinases.
| Signal protein | Receptor | Activity |
|---|---|---|
| Epidermal growth factor | EGF‐R | Stimulates cell growth and differentiation |
| Insulin | Insulin‐R | Enhances glucose consumption and protein synthesis |
| Insulin‐like growth factor | IGF‐1‐R | Stimulates cell growth and survival in many cell types |
| Nerve growth factor (NGF) | Trk R | Stimulates cell growth and survival of neurons |
| Platelet‐derived growth factor (PDGF) | PDGF‐R | Stimulates cell growth, differentiation and cell migration |
| Macrophage colony‐stimulating factor (MCSF) | MCSF‐R | Stimulates cell growth and differentiation of macrophages and monocytes |
| Fibroblast growth factors (FGF1–FGF24) | FGF‐R | Stimulates cell growth and differentiation |
| Vascular endothelial growth factor (VEGF) | VEGF‐R | Stimulates angiogenesis |
| Ephrin | Eph‐R | Stimulates angiogenesis and axon orientation |
R, receptor.
These signal pathways have in common that they amplify the original signal from a few signal molecules to thousands of second messengers(cAMP, Ca 2+), which can trigger thousands of targets ( Figure 3.11). The pathways downstream the original receptor are often not linear but complex networks. Members of pathways and networks often assume roles in more than a single context, which make their analysis far from easy. Many elements of these pathways behave like molecular switches, which can quickly change their state from active to inactive. Reversible phosphorylation/dephosphorylation and binding of GTP/GDP or ATP/ADP are common elements of these switches ( Figure 2.16). Many pathways and networks are apparently regulated by positive and negative feedback mechanism.
3.1.2 Endomembrane System in a Eukaryotic Cell
Most eukaryotes have an extensive endomembrane systemcovering the entire intracellular space. The most striking parts are the ERand the Golgi complex. The ER is especially elaborated in endocrine cells. Other compartments are also contained in biomembranes and form separate reaction entities within the cell. The characteristics of internal biomembranes may vary, depending on the membrane proteins and lipids they contain.
The ER ( Figure 3.12) is an extensive labyrinth of tubules and sacs pervading the entire eukaryotic cell. It is here that the components of biomembranes are assembled, and it is here that the posttranslational modificationof proteins takes place. The rough ER contains ribosomes, which produce (translate) proteins ready for export (see Section 5.3), while the smooth ERis ribosome‐free and contains enzymes requiring a lipophilic environment, such as cytochrome oxidases.
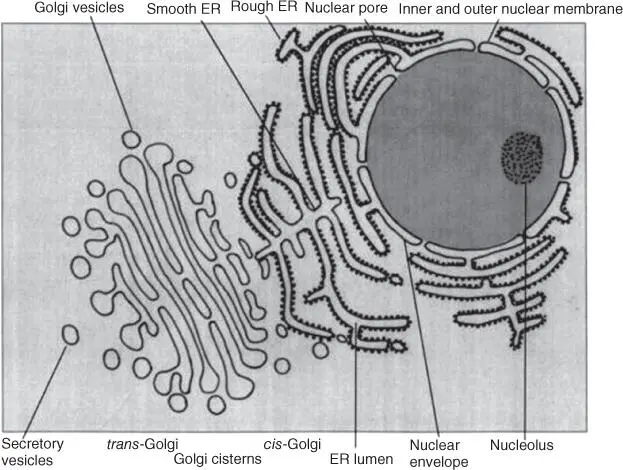
Figure 3.12 Schematic representation of the endomembrane system of the cell: nuclear envelope, rough and smooth endoplasmic reticulum (ER), and Golgi complex.
The ER envelopes the eukaryotic nucleus, which is thus surrounded by two biomembranes ( Figure 3.12). The nuclear membrane has two characteristic nuclear pore complexes regulating the entry of molecules (e.g. transcription factors and ribosomal proteins) into the nucleus as well as the export (e.g. of mRNA and ribosomal subunits) (see Section 5.1). The nucleus is one of the most striking and characteristic organelles in a eukaryotic cell, containing genetic information, stored as DNA. DNA is usually found in several linear double strands or chromosomes, as seen under the light microscope. It is usually surrounded by proteins (e.g. histones) that form specific structures known as nucleosomes(see Section 4.1.2). The nucleolus, a distinct structure visible under the electron microscope, contains rRNAs serving as scaffolding for the assembly of ribosomal subunits.
Piles of membranous tubules form the Golgi complex( Figure 3.12). The Golgi complex receives vesicles containing proteins from the ER on the cis side and passes them on for transport on the trans side to lysosomesor to the cytoplasmic membrane for export. The proteins are modified in the Golgi complex – sugar residues are cleaved off or added (see Sections 5.3and 5.4). The Golgi complex is particularly developed in glandular cells.
Lysosomes( Figure 3.13) are small membrane‐enclosed organelles with an irregular structure. They contain a range of hydrolytic enzymes ( nucleases, proteases, glycosidases, lipases, phosphatases, sulfatases, and phospholipases) that degrade lipids, polysaccharides, proteins, and nucleic acids. Liposomes also degrade and recycledefective macromolecules or organelles. Monomers released from proteins, polysaccharides, and lipids are often recyclable. Lysosomes evolve from vesicles cut off from the Golgi complex, also known as endosomes. Their pH value is acidic, due to membrane‐bound H +‐ATPases pumping protons into the lysosomes. Hydrolytic enzymes have a pH optimum of 4–5 and become inactive at pH 7. Thus, should any of the hydrolytic enzymes escape into the cytoplasm, which has a pH around 7.4, they cannot cause any harm. Lysosomesfuse with endosomesor phagosomesthat are pinched off from the cytoplasmic membrane by endocytosis and filled with protein complexes or microorganisms (see Section 5.4).
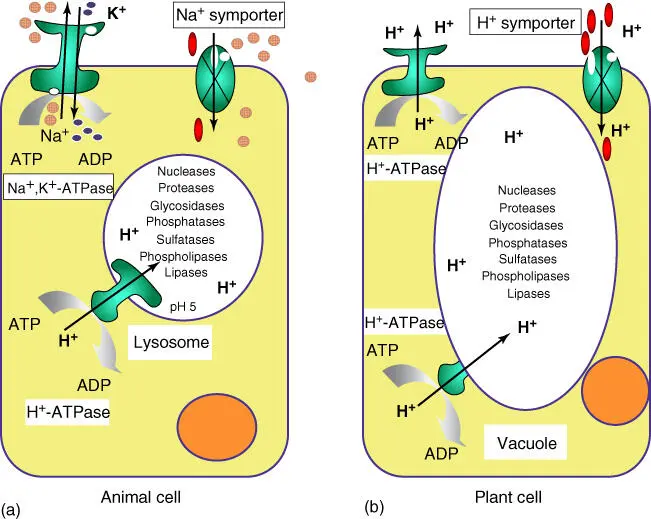
Figure 3.13 Similarities of lysosomes and plant vacuoles. (a) Schematic structure of lysosomes. (b) Schematic structure of plant cells with vacuoles.
Plant cellsdo not contain lysosomes, but vacuoles. These can make up by far the largest compartments in adult plant cells (Figures 1.2and 3.13). Vacuoles store inorganic ions and low‐molecular‐weight metabolites (e.g. sugar, organic acids, and amino acids). All plants produce secondary compoundssuch as flavonoids, phenylpropans, tannins, terpenes, iridoid glycosides, alkaloids, glucosinolates, and cyanogenic glycosides, which are not needed for their primary metabolism. Contrary to earlier beliefs, they are not waste products, but ensure the survival of the plant, defending it against herbivores and microorganisms. As signal compounds, they can also help communicate with other organisms by attracting insects for pollination or animals for seed propagation. Polar secondary compoundsare frequently stored in vacuoles, whereas lipophilic compoundsare kept in oil vessels, resin channels, or glandular cells. Often, secondary compounds are stored as prodrugs, and only when the plant is wounded or an infection occurs will they be activated – mostly by β‐glucosidasecleaving off a glucose residue. Seeds contain vacuoles storing a protein reserve. Vacuoles can have various functions in a plant cell – providing additional storage spaceor acting as a defense or signaling compartment. The storage function in particular requires high osmotic pressure inside the vacuole, which is crucial for the stabilization and growth of the plant ( turgor regulation). Active transport in plants mostly functions via proton gradients, as opposed to animal cells, which rely more on Na +/K +gradients, using Na +, K +‐ATPase (see Section 3.1.1.2).
Читать дальшеИнтервал:
Закладка:
Похожие книги на «An Introduction to Molecular Biotechnology»
Представляем Вашему вниманию похожие книги на «An Introduction to Molecular Biotechnology» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.


![Andrew Radford - Linguistics An Introduction [Second Edition]](/books/397851/andrew-radford-linguistics-an-introduction-second-thumb.webp)









Обсуждение, отзывы о книге «An Introduction to Molecular Biotechnology» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.