John Long - Darwin’s Devices
Здесь есть возможность читать онлайн «John Long - Darwin’s Devices» весь текст электронной книги совершенно бесплатно (целиком полную версию без сокращений). В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: Биология, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Darwin’s Devices
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Darwin’s Devices: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Darwin’s Devices»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
In Darwin’s Devices, Long tells the story of these evolving biorobots—how they came to be, and what they can teach us about the biology of living and extinct species. Evolving biorobots can replicate creatures that disappeared from the earth long ago, showing us in real time what happens in the face of unexpected environmental challenges. Biomechanically correct models of backbones functioning as part of an autonomous robot, for example, can help us understand why the first vertebrates evolved them.
But the most impressive feature of these robots, as Long shows, is their ability to illustrate the power of evolution to solve difficult technological challenges autonomously—without human input regarding what a workable solution might be. Even a simple robot can create complex behavior, often learning or evolving greater intelligence than humans could possibly program. This remarkable idea could forever alter the face of engineering, design, and even warfare.
An amazing tour through the workings of a fertile mind, Darwin’s Devices will make you rethink everything you thought you knew about evolution, robot intelligence, and life itself.
Darwin’s Devices — читать онлайн бесплатно полную книгу (весь текст) целиком
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Darwin’s Devices», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
We had three reasons for thinking that Tadro3 was the Evolvabot we were looking for: (1) the brain would make it autonomous, able to behave on its own without a human “in the loop,” without a remote operator acting as the eyes and brains of the operation; (2) the light-seeking behavior would emulate the phototaxis of the tunicate tadpole larva; and (3) the body would also emulate that of the tunicate larva, possessing a propulsive tail with a biomimetic notochord, a backbone whose properties we could vary by degrees, code with an artificial genome, and cause to evolve under the right ecological situation.
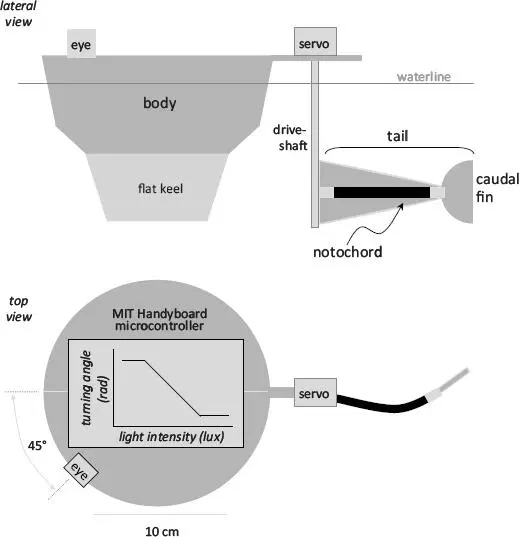
FIGURE 3.3. Tadro3, the Evolvabot designed to represent the tunicate tadpole larva. Tadro3 has a single eyespot (photoresistor), a flapping tail, and a microcontroller that converts the light intensity at the eyespot into a turning angle at the tail. This sensorimotor system produces autonomous phototactic navigation (see Figure 3.2). Tadro3 has a biomimetic gelatin hydrogel serving as a notochord. The notochord’s structural stiffness is determined by the material stiffness of the gelatin, which we control with chemical cross-linking, and the length of the tail. Both material stiffness and length of the tail were coded genetically as evolvable characters. Proportions are drawn to scale, and more information about the specifications of the design are available. [20] J. H. Long Jr., T. J. Koob, K. Irving, K. Combie, V. Engel, N. Livingston, A. Lammert, and J. Schumacher, “Biomimetic Evolutionary Analysis: Testing the Adaptive Value of Vertebrate Tail Stiffness in Autonomous Swimming Robots,” Journal of Experimental Biology 209, no. 23 (2006): 4732–4746.
To understand more about the backbone as a feature we targeted, we need to dig a bit deeper into some of the assumptions we’ve been making about the evolution of notochords and vertebral columns of chordates. As I said in Chapter 1, a species called Haikouichthys ercaicunensis , a small, sporty little fish that lived 530 million years ago (Figure 3.2), appears to have been conducting its own evolutionary experiment on turning a notochord into a vertebral column. Widely spaced bits of cartilage or bone can be seen along its notochord. [21] You can see the original pictures of the fossils in this paper: D.-G. Shu, S. Conway Morris, J. Han, Z.-F. Zhang, K. Yasui, P. Janvier, L. Chen, X.-L. Zhang, J.-N. Liu, Y. Li, and H.-Q. Lui, “Head and Backbone of the Early Cambrian Vertebrate Haikouichthys ,” Nature 421, no. 6922 (January 2003): 526–529.
The proto-vertebrae, as some authors have dubbed them, are too far apart to resemble the tightly packed vertebrae that we see in most other fossils or living species that have vertebrae. However, for all of their differences, the proto-vertebrae of Haikouichthys allow us to infer three important things about the evolution of vertebrae:
* The earliest vertebrate fossils had a backbone that was primarily a notochord, supporting the contention that the notochord is the an cestral state of the vertebrate axial skeleton (no one is surprised by this, by the way, because evolutionary trees have long inferred this pattern, as we’ll see in a minute).
* Vertebrae, even though they appear early in vertebrate evolution, take millions of years to evolve into what we now recognize as a vertebral column. Philippe Janvier, a paleontologist specializing in the earliest fishes, estimates the origin of an internal skeleton of calcified cartilage or bone at about 443 million years ago, about 90 million years after Haikouichthys ’ experiments.
* Because the backbone of Haikouichthys does not have the large vertebrae and thin intervertebral joints that we see in living fishes, but just the opposite, we need to be careful to recognize that the two states of the axial skeleton, notochords and vertebral columns, really demarcate the ends of a spectrum of possible axial skeletons. With that in mind, we’d expect to see throughout living and extinct vertebrates variations in the size, shape, and number of vertebrae and intervertebral joints.
Phylogenetic analysis gives us another clue about the polarity of the states, or spectrum of states, of the axial skeleton. The notochord, without any signs of vertebrae, is possessed by both tunicates and lancelets (see Figure 3.2). If, as Delsuc’s tree showed, tunicates are the sister group to vertebrates and lancelets are the sister group to tunicates + vertebrates, then the simplest, most parsimonious explanation is that notochords evolved in the common ancestor of all three groups, well before the vertebrates split off and began to evolve the vertebrae that we think we see in Haikouichthys.
Additional evidence for the notochord evolving first is that it also appears first in the development of living fish, prior to the formation of vertebrae; vertebrae are then built in and around the notochord. [22] Sindre Grotmol, Harald Hryvi, Roger Keynes, Christel Krossøy, Kari Nordvik, and Geir K. Totland, “Stepwise Enforcement of the Notochord and Its Intersection with the Myoseptum: An Evolutionary Path Leading to Development of the Vertebra?” Journal of Anatomy 209, no. 3 (2006): 339–357.
Although being first in development isn’t, by itself, evidence for evolutionary polarity, the notochord is a central structure in early embryo development, one that is necessary for the formation of the nervous system and the growth of the embryo. Every vertebrate embryo grows a notochord first and then, if they grow one at all, a vertebral column. This invariant pattern of the notochord guiding the embryonic development of vertebrates and their vertebrae is consistent with the hypothesis that notochords evolved before vertebral columns.
In development and evolution the axial skeleton functions to stiffen the body. As we talked about in Chapter 1, stiffness is the mechanical property that dictates how much a structure changes shape—lengthens, shortens, twists, or bends—in response to having forces applied to it. Put a rubber band on your two index fingers and apply a tensile force to it by increasing the distance between your fingers. The rubber band, at least at first, lengthens easily. Now do the same thing with a shoelace, the ends of which you hold between index finger and thumb. The shoelace does not lengthen much, even if you apply as much force as you can. In engineering terms the shoelace is “stiffer in tension” than the rubber band.
Bending or flexural stiffness of the notochord can be increased by adding vertebrae. [23] “Stiffness” by itself is an ambiguous term. Many different kinds of stiffness exist. What they have in common is that they are all a proportionality constant between an applied force, stress, or torque and the resulting change in length, strain, or curvature, respectively. Flexural stiffness is defined as the proportionality constant between a torque and the resulting curvature. Flexural stiffness assumes that this relationship is the same all along the length of the structure that you are bending. If you want to describe the stiffness of the whole structure, a physicist would talk about “spring constant” or a “spring stiffness.” I prefer the term “structural stiffness.” In the case of a cantilevered beam with a weight hung off the end, the structural stiffness is the ratio of the flexural stiffness to the cube of the beam’s length. This means that you can have beams of constant flexural stiffness but different structural stiffness if you vary their lengths.
Working with Tom Koob and Lena Koob-Emunds at the Mount Desert Island Biological Laboratory in Salsbury Cove, Maine, we analyzed hagfish, a group of eel-like fish that never evolved jaws and retain, as adults, a fifty-centimeter long notochord. After a hagfish died, we removed and bent its notochord to measure the notochord’s flexural stiffness. We then threaded onto the notochord, like pearls on a string, a series of rigid plastic rings that snugly fit the notochord. Sometimes we added just a few rings, widely spaced like the vertebrae of Haikouichthys, and sometimes we added more, leaving less space for bending. The result? More vertebrae created an axial skeleton with increased flexural stiffness. With this in mind, in Tadro3 we allowed bending stiffness itself, rather than number of vertebrae, to be the character that was genetically coded to evolve.
Интервал:
Закладка:
Похожие книги на «Darwin’s Devices»
Представляем Вашему вниманию похожие книги на «Darwin’s Devices» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Darwin’s Devices» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.