Jane Flint - Principles of Virology
Здесь есть возможность читать онлайн «Jane Flint - Principles of Virology» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.
- Название:Principles of Virology
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Principles of Virology: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Principles of Virology»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Volume I: Molecular Biology
Volume II: Pathogenesis and Control
Principles of Virology, Fifth Edition
Principles of Virology — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Principles of Virology», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
A protein platform also appears to participate in de novo priming by the reovirus RdRP, a cube-like structure with a catalytic site in the center that is accessible by four tunnels. One tunnel allows template entry, one serves for the exit of newly synthesized double-stranded RNA, a third permits exit of mRNA, and a fourth is for substrate entry. A priming loop that is not observed in this region of other RNA polymerases is present in the palm domain. The loop supports the initiating NTP, then retracts into the palm and fits into the minor groove of the double-stranded RNA product. This movement assists in the transition between initiation and elongation, and also allows the newly synthesized RNA to exit the polymerase.
Protein platforms also appear to participate in the de novo priming of RNA synthesis by flaviviruses other than hepatitis C virus (dengue and West Nile viruses), influenza virus genome RNA synthesis, all known (–) strand RNA viruses, and bacteriophage Φ6.
Primer-Dependent Initiation
Protein priming.A protein-linked oligonucleotide serves as a primer for RNA synthesis by RdRPs of members of the Picornaviridae and Caliciviridae . Protein priming also occurs during DNA replication of adenoviruses, certain DNA-containing bacteriophages ( Chapter 10), and hepatitis B virus ( Chapter 7). A terminal protein provides a hydroxyl group (in a tyrosine or serine residue) to which the first (priming) oligonucleotide can be linked by viral polymerases, via a phosphodiester bond. The primer is then elongated.
Polioviral genomic RNA, as well as newly synthesized (+) and (−) strand RNAs, are covalently linked at their 5′ ends to the 22-amino-acid protein VPg ( Fig. 6.9A), initially suggesting that VPg might function as a primer for RNA synthesis. This hypothesis was supported by the discovery of a uridylylated form of the protein, VPg-pUpU, in infected cells. VPg can be uridylylated in vitro by 3D poland can then prime the synthesis of VPg-linked poly(U) from a poly(A) template. The template for uridylylation of VPg is either the 3′ poly(A) on (+) strand RNA [during synthesis of (−) strand RNA] ( Fig. 6.10) or an RNA hairpin, the cis -acting replication element (cre), located in the coding region [during synthesis of (+) strand RNA] ( Fig. 6.9B and C).
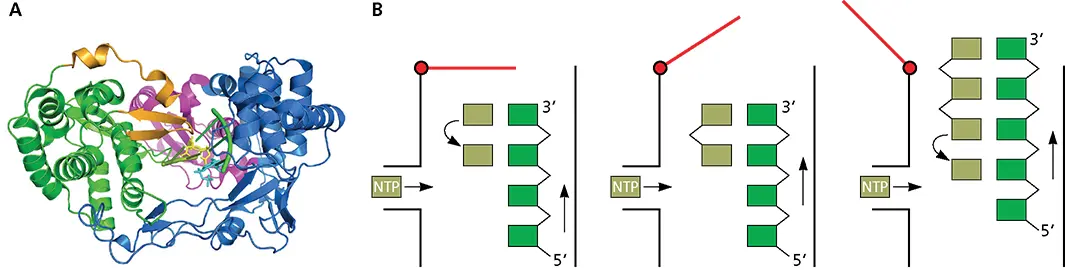
Figure 6.8 Mechanism of de novo initiation. (A)Ribbon diagram of RdRP of hepatitis C virus. Fingers, palm, and thumb domain are colored blue, green, and magenta, respectively. The C-terminal loop that blocks the active site is shown in brown. Active-site residues are yellow (PDB file 4WTM). (B)Swinging-gate model of initiation. With the RNA template (green) in the active site of the enzyme, a short β-loop (red) provides a platform on which the first complementary nucleotide (light green) is added to the template (left). The second nucleotide is then added, producing a dinucleotide primer for RNA synthesis (middle). At this point, nothing further can happen because the priming platform blocks the exit of the RNA product from the enzyme. The solution to this problem is that the polymerase undergoes a conformational change that moves the priming platform out of the way and allows the newly synthesized complementary RNA (right) to exit as the enzyme moves along the template strand.
Structures of the RdRPs of different picornaviruses and caliciviruses indicate that the active site is more accessible than in polymerases with a de novo mechanism of initiation. The small thumb domains of these polymerases leave a wide central cavity that can accommodate the template and the protein primer.
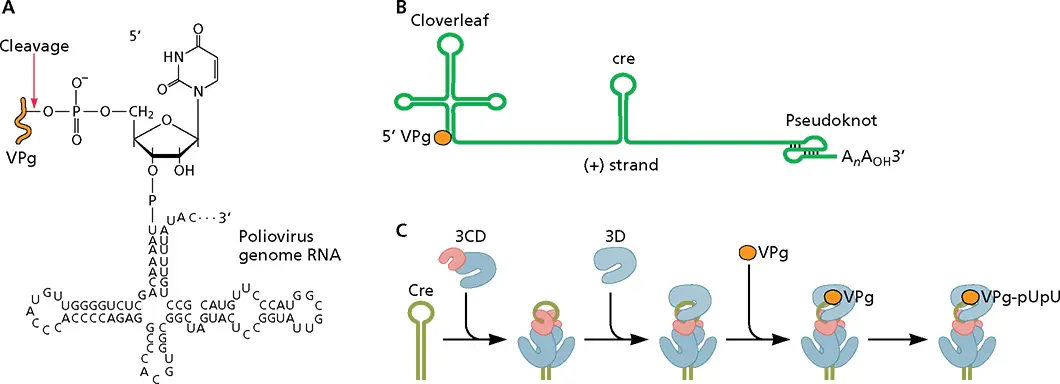
Figure 6.9 Uridylylation of VPg. (A)Linkage of VPg to polioviral genomic RNA. Polioviral RNA is linked to the 22-amino-acid VPg (orange) via an O 4-(5′-uridylyl)-tyrosine linkage. This phosphodiester bond is cleaved at the indicated site by a cellular enzyme to produce the viral mRNA containing a 5′-terminal pU. (B)Structure of the poliovirus (+) strand RNA template, showing the 5′ cloverleaf structure, the internal cre ( cis -acting replication element) sequence, and the 3′ pseudoknot. (C)Model for assembly of the VPg uridylylation complex. Two molecules of 3CD bind to cre. The 3C dimer melts part of the stem. 3D polbinds to the complex by interactions between the back of the thumb domain and the surface of 3C. VPg then binds the complex and is linked to two U moieties in a reaction templated by the cre sequence.
Biochemical and structural studies have identified three different VPg binding sites on 3D pol. Uridylylation of foot- and-mouth disease virus VPg can be achieved in a reaction containing 3D pol, a template (rA) 10, UTP, and Mg 2+and Mn 2+. Crystallographic analysis of 3D polcarrying out uridylyation reveals that VPg-pU is bound in the template-binding channel, with the N terminus of VPg in the NTP entry channel and the C terminus pointing toward the template-binding channel. The hydroxyl group of a tyrosine in VPg is covalently linked to the α-phosphate of UMP and interacts with a divalent metal ion that binds an Asp of the Gly-Asp-Asp motif in the active site. This arrangement of VPg is similar to that of the primer terminus in the nucleotidyl transfer reaction, demonstrating that 3D polcatalyzes VPg uridylylation using the same two-metal mechanism as the nucleotidyl transfer reaction. A second binding site for VPg has been located on the base of the thumb subdomain of the polymerase of Coxsackievirus B3, in a position that does not allow uridylylation in cis . It has been suggested that uridylylation of VPg might be accomplished in trans by another 3D polmolecule. A third binding site on the 3D polof enterovirus 71 is at the base of the palm and also would require uridylylation by another polymerase molecule.
When VPg uridylylation begins at the 3′-poly(A) tail of the (+) strand template, the polymerase continues nucleotidyl transfer reactions and copies the entire genome. However, when uridylylation of VPg takes place on the cre, the protein must dissociate and transfer to the 3′ end of the RNA. How this process is accomplished is not known ( Fig. 6.10).
Protein priming by the birnavirus RdRP VP1 is unusual because the primer isthe polymerase, not a separate protein. Even in the absence of a template, VP1 has self-guanylylation activity that is dependent on divalent metal ions. The guanylylation site is a serine located approximately 23 Å from the catalytic site of the polymerase. The long distance between these sites suggests that guanylylation may be carried out at a second active site. The finding that some altered polymerases that are inactive in RNA synthesis retain self-guanylylation activity supports this hypothesis. After two G residues are added to VP1, it binds to a conserved CC sequence at the terminus of the viral RNA template to initiate RNA synthesis. The 5′ ends of mRNAs and genomic double-stranded RNAs produced by this reaction are therefore linked to a VP1 molecule.
Priming by capped RNA fragments.Influenza virus mRNA synthesis is blocked by treatment of cells with the fungal toxin α-amanitin at concentrations that inhibit cellular DNA-dependent RNA polymerase II. This surprising finding demonstrated that the viral RNA polymerase is dependent on host cell RNA polymerase II. Inhibition by α-amanitin is explained by a requirement for newly synthesized cellular transcripts made by this enzyme to provide primers for influenza viral mRNA synthesis ( Fig. 6.11). Presumably, these cellular transcripts must be made continuously because they are exported rapidly from the nucleus once processed. Such transcripts are cleaved in the nucleus by an influenza virus-encoded, cap-dependent endonuclease that is part of the RdRP ( Fig. 6.12). The resulting 10- to 13-nucleotide capped fragments serve as primers for the initiation of viral mRNA synthesis.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Principles of Virology»
Представляем Вашему вниманию похожие книги на «Principles of Virology» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Principles of Virology» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.