Jane Flint - Principles of Virology
Здесь есть возможность читать онлайн «Jane Flint - Principles of Virology» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.
- Название:Principles of Virology
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Principles of Virology: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Principles of Virology»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Volume I: Molecular Biology
Volume II: Pathogenesis and Control
Principles of Virology, Fifth Edition
Principles of Virology — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Principles of Virology», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Closure of the active site by movements of the palm domain appears to be a feature of the RdRPs of all (+) RNA viruses but not (–) strand or double-stranded RNA viruses; their palm domains are already structured in the unbound form. This simple nucleotide selection mechanism greatly influences polymerase fidelity. In T7 RNA polymerase and Taq DNA polymerase, a pre-insertion site is utilized to first bind the incoming NTP to the templating base. Next, the template-NTP base pair undergoes a major movement that places the triphosphate into the active site. These higher-fidelity enzymes therefore select for proper template-NTP pairing at two different binding sites, in contrast to the single site used by (+) strand RNA virus RdRPs.
Functions of Additional Polymerase Domains
Additional N- and C-terminal domains that surround the RdRP cores are often encoded in the genomes of larger RNA viruses ( Fig. 6.13). The flavivirus RdRP has an extra N-terminal domain that has 5′-methyltransferase activity that contributes to mRNA capping. The core RdRPs of double-stranded RNA viruses are flanked by large N- and C-terminal domains. The former surrounds the fingers and thumb subdomains, closing the enzyme in a cage-like structure. The C-terminal domains are shaped like bracelets and resemble the sliding clamps that contribute to the efficiency of DNA polymerases. In contrast to other RdRPs, these enzymes have four channels. Two are in equivalent positions to the template and nucleotide entry channels of other RdRPs, but the other two serve as RNA exit pathways. One extends through the bracelet domain and is the pathway for release of new double-stranded RNA to the particle interior. The other serves to guide newly synthesized (+) single-stranded RNAs out of the core.
The RdRP of (–) strand RNA bunyaviruses has an N-terminal endonuclease domain that is essential for procuring capped mRNA primers. The minimal RdRP of vesicular stomatitis virus is surrounded by three globular domains with three enzymatic activities required for mRNA 5′-cap synthesis: 2′- O -methyltransferase, guanine-N7-methyltransferase, and polyribonucleotidyl transferase.
Not all RdRPs have other functions encoded in extra N- and C-terminal domains. The influenza virus RdRP consists of three individual polypeptides, PA, PB1, and PB2, each of which has the distinct activity described above.
RNA Polymerase Oligomerization
RNA polymerases of multiple (+) and (–) strand RNA viruses have been reported to form dimers and higher-order oligomers ( Fig. 6.14). There is evidence that such arrangements may increase the stability and catalytic activity of these enzymes. In many cases, deletions of amino acids that prevent oligomerization also inhibit or cause complete loss of enzyme activity.
The first poliovirus 3D polstructure revealed that the polymerase molecules interacted in a head-to-tail manner and formed fibers; subsequently the protein was shown to form a lattice. The head-to-tail fibers were formed by an interface comprising parts of the thumb of one polymerase and the back of the palm of another. Amino acid changes in the back of the thumb that disrupt this interface impaired replication. Repetition of this interaction in a head-to-tail fashion results in long fibers of polymerase molecules 50 Å in diameter. The presence of a second interface, formed by N-terminal polypeptide segments, may lead to a network of polymerase fibers. These interacting N-terminal polypeptide segments may originate from different polymerase molecules and are required for enzyme activity. Intermolecular cross-linking has been observed between cysteines engineered at Ala29 and Ile441 of poliovirus 3D pol, and disruption of these interactions led to reduced infectivity. Polymerase-containing oligomeric structures resembling those seen with purified 3D polwere observed on the surface of vesicles isolated from poliovirus-infected cells. Because picornavirus RNA synthesis occurs on membranous vesicles, the concept of a catalytic lattice is attractive mechanistically.
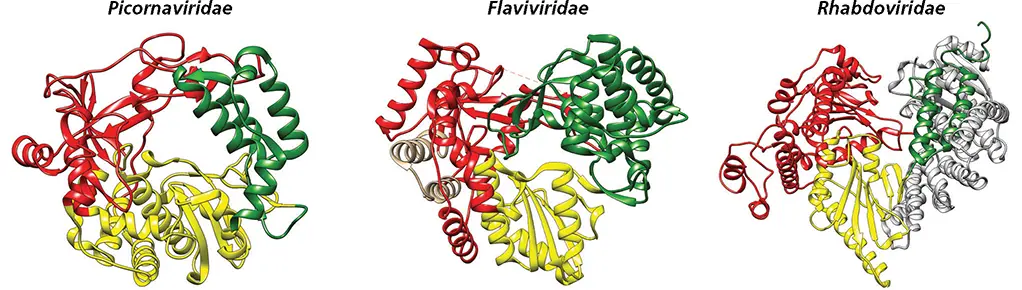
Figure 6.13 Functional N- and C-terminal extensions of RNA polymerases. The smallest known RdRP is encoded in picornavirus genomes and consists of a core catalytic unit made of thumb (green), fingers (red), and palm (yellow) domains. An N-terminal extension of the flavivirus dengue virus RNA polymerase (tan) has methyltransferase activity. The rhabdovirus RNA polymerase has both N- (blue) and C-terminal (light gray) extensions; the latter contain the capping and methyltransferase domains.
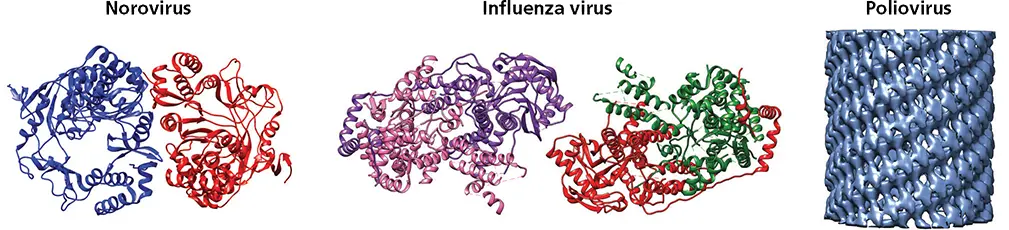
Figure 6.14 Oligomerization of RNA-dependent RNA polymerases. Ribbon diagrams of dimers of murine norovirus RdRP (PDB ID: 3QID), tetramers of influenza virus RdRP (PDB ID: 3J9B), and cryo-electron microscopy density data showing tubular arrangement of sheets of poliovirus 3D pol(EMD ID: 2270).
Template Specificity
Viral RdRPs must select viral templates from among a vast excess of cellular mRNAs and then initiate correctly to ensure accurate RNA synthesis. Different mechanisms that contribute to template specificity have been identified. Initiation specificity may be regulated by the affinity of the RdRP for the initiating nucleotide. For example, the RdRPs of bovine viral diarrhea virus and bacteriophage ϕ6 prefer 3′-terminal C. Reovirus RdRP prefers a G at the second position of the template RNA. This preference is controlled by hydrogen bonding of carbonyl and amino groups of the G with two amino acids of the enzyme. Both preferences would exclude initiation on cellular mRNAs, the great majority of which end in poly(A).
Template specificity may also be conferred by the recognition of RNA sequences or structures at the 5′ and 3′ ends of viral RNAs by viral proteins. RNA synthesis initiates specifically within a polypyrimidine tract in the 3′ untranslated region of hepatitis C virus RNA. The 3′ noncoding region of polioviral genomic RNA contains an RNA pseudoknotstructure that is conserved among picornaviruses ( Fig. 6.9). A viral protein (3AB-3CD) binds this structure and may direct the RdRP to that site for the initiation of (−) strand RNA synthesis. The precursor to poliovirus 3CD proplays an important role in viral RNA synthesis by participating in the formation of a ribonucleoprotein at the 5′ end of the (+) strand RNA. This protein, together with cellular poly(rC)-binding protein 2, binds to a cloverleaf structure in the viral RNA ( Fig. 6.9). Alterations within the RNA-binding domain of 3CD inhibit binding to the cloverleaf and RNA synthesis.
Internal RNA sequences may also confer initiation specificity to RdRPs. The cis -acting replication elements (cre) in the coding sequence of poliovirus protein 2C and rhinovirus capsid protein VP1 contain short RNA sequences that are required for RNA synthesis. These sequences are binding sites for 3CD proand, as discussed previously, serve as a template for uridylylation of the VPg protein ( Fig. 6.9).
During mRNA synthesis by influenza virus polymerase, sequences at the RNA termini ensure that the 5′ ends of newly synthesized viral mRNAs are not cleaved and used as primers ( Fig. 6.12). If such cleavage were to occur, there would be no net synthesis of viral mRNAs. Polymerase binding to two sites in the genomic RNA blocks access of a second P protein and protects newly synthesized viral mRNA from endonucleolytic cleavage by P proteins.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Principles of Virology»
Представляем Вашему вниманию похожие книги на «Principles of Virology» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Principles of Virology» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.