George Acquaah - Principles of Plant Genetics and Breeding
Здесь есть возможность читать онлайн «George Acquaah - Principles of Plant Genetics and Breeding» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Principles of Plant Genetics and Breeding
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Principles of Plant Genetics and Breeding: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Principles of Plant Genetics and Breeding»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Organizes topics to reflect the stages of an actual breeding project Incorporates the most recent technologies in the field, such as CRSPR genome edition and grafting on GM stock Includes numerous illustrations and end-of-chapter self-assessment questions, key references, suggested readings, and links to relevant websites Features a companion website containing additional artwork and instructor resources
offers researchers and professionals an invaluable resource and remains the ideal textbook for advanced undergraduates and graduates in plant science, particularly those studying plant breeding, biotechnology, and genetics.
Principles of Plant Genetics and Breeding — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Principles of Plant Genetics and Breeding», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Self‐incompatibility systems
Self‐incompatibility systems may be classified into two basic types: heteromorphicand homomorphic.
Heteromorphic incompatibilityThis is caused by differences in the lengths of stamens and style (called heterostyly) ( Figure 5.5). In one flower type called the pin, the styles are long while the anthers are short. In the other flower type, thrum, the reverse is true (e.g. in Primula). The pin trait is conditioned by the genotype ss while thrum is conditioned by the genotype Ss. A cross of pin (ss) × pin (ss) as well as thrum (Ss) × thrum (Ss) are incompatible. However, pin (ss) × thrum (Ss), or vice versa, is compatible. The condition described is distyly because of the two different types of style lengths of the flowers. In Lythrum three different relative positions occur (called tristyly).
Homomorphic incompatibilityThere are two kinds of homomorphic incompatibility: gametophytic and sporophytic ( Figure 5.6).Gametophytic incompatibilityIn gametophytic incompatibility (originally called the oppositional factor system), the ability of the pollen to function is determined by its own genotype and not the plant that produces it. Gametophytic incompatibility is more widespread than sporophytic incompatibility. Gametophytic incompatibility occurs in species such as red clover, white clover, and yellow sweet clover. Homomorphic incompatibility is controlled by a series of alleles at a single locus (S1, S2, …, Sn) or alleles at two loci in some species. The system is called homomorphic because the flowering structures in both the seed‐bearing (female) and pollen‐bearing (male) plants are similar. The alleles of the incompatibility gene(s) act individually in the style. They exhibit no dominance. The incompatible pollen is inhibited in the style. The pistil is diploid and hence contains two incompatibility alleles (e.g. S1S3, S3, S4). Reactions occur if identical alleles in both pollen and style are encountered. Only heterozygotes for S alleles are produced in this system.Sporophytic incompatibilityIn sporophytic incompatibility, the incompatibility characteristics of the pollen are determined by the plant (sporophyte) that produces it. It occurs in species such as broccoli, radish, and kale. The sporophytic system differs from the gametophytic system in that the S allele exhibits dominance. Also, it may have individual action in both pollen and style, making this incompatibility system complex. The dominance is determined by the pollen parent. Incompatible pollen may be inhibited on the stigma surface. For example, a plant with genotype S1S2 where S1 is dominant to S2, will produce pollen that will function like S1. Furthermore, S1 pollen will be rejected by an S1 style but received by an S2 style. Hence, homozygotes of S alleles are possible.Incompatibility is expressed in one of three general ways, depending on the species. The germination of the pollen may be decreased (e.g. in broccoli). Sometimes, removing the stigma allows normal pollen germination. In the second way, pollen germination is normal, but pollen tube growth is inhibited in the style (e.g. tobacco). In the third scenario, the incompatibility reaction occurs after fertilization (e.g. in Gesteria). This third mechanism is rare.
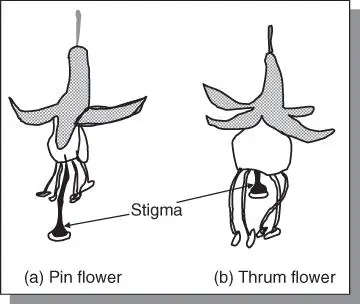
Figure 5.5Heteromorphic incompatibility showing floral modifications in which anthers and pistils are of different lengths in different plants (heterostyly). This type of incompatibility is believed to be always of the sporophytic type. Pin and thrum flowers occur in flowers such as Primula, Forsythia, Oxalis, and Silia .
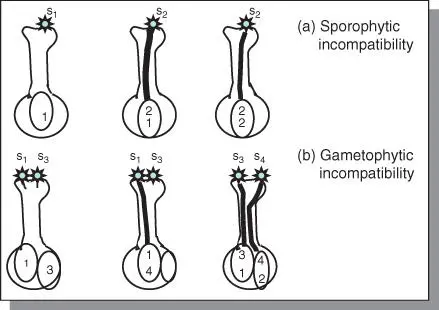
Figure 5.6Types of self‐incompatibility: (a) sporophytic, and (b) gametophytic. Sporophytic incompatibility occurs in families such as Compositae and Cruciferae. It is associated with pollen grains with two generative nuclei, whereas gametophytic incompatibility is associated with pollen with one generative nucleus in the pollen tube as occurs in various kinds of clover.
Changing the incompatibility reaction
Mutagens (agents of mutation) such as X‐rays, radioactive sources such as P 32, and certain chemicals have been used to make a self‐infertile genotype self‐fertile. Such a change is easier to achieve in gametophytic systems than sporophytic systems. Furthermore, doubling the chromosome number of species with the sporophytic system of incompatibility does not significantly alter the incompatibility reaction. This is because two different alleles already exist in a diploid that may interact to produce the incompatibility effect. Polyploidy only makes more of such alleles available. On the other hand, doubling the chromosome in a gametophytic system would allow the pollen grain to carry two different alleles (instead of one). The allelic interaction could cancel any incompatibility effect to allow selfing to be possible. For example, diploid pear is self‐incompatible whereas autotetraploid pear is self‐fruitful.
Plant breeding implications of self‐incompatibility
Infertility of any kind hinders plant breeding. However, this handicap may be used as a tool to facilitate breeding by certain methods. Self‐incompatibility may be temporarily overcome by techniques or strategies such as the removal of the stigma surface ( Figure 5.7) (or application of electric shock), early pollination (before inhibitory proteins form), or lowering the temperature (to slow down the development of the inhibitory substance). Self‐incompatibility promotes heterozygosity. Consequently, selfing self‐incompatible plants can create significant variability from which a breeder can select superior recombinants. Self‐incompatibility may be used in plant breeding (for F 1hybrids, synthetics, triploids), but first homozygous lines must be developed.

Figure 5.7Cut‐style and embryo rescue in lily. Overcoming reproductive barriers. (a) Pollination barriers; (b) post‐fertilization reproductive barriers. Courtesy of JM van Tuyl/P Ariens Wageningen UR ‐ Plant Breeding.
Self‐incompatibility systems for hybrid seed production have been established for certain crops (e.g. cabbage, kale) that exhibit sporophytic incompatibility ( Figure 5.8). Inbred lines (compatible inbreds) are used as parents. These systems generally are used to manage pollinations for commercial production of hybrid seed. Gametophytic incompatibility occurs in vegetatively propagated species. The clones to be hybridized are planted in adjacent rows.
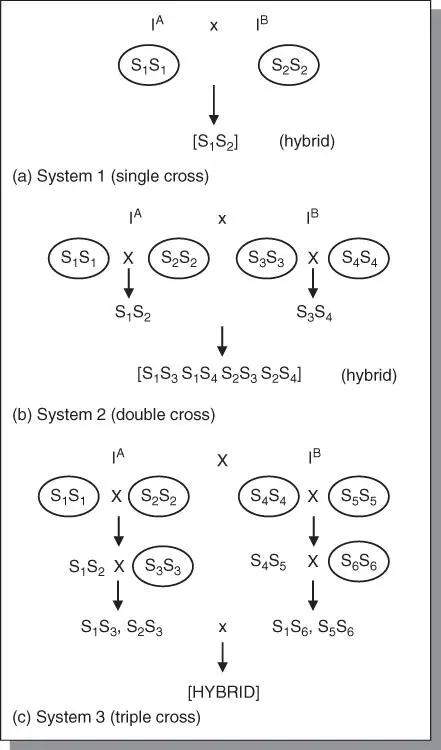
Figure 5.8Application of self‐incompatibility in practical plant breeding. Sporophytic incompatibility is widely used in breeding of cabbage and other Brassica species. The single cross hybrids are more uniforms and easier to produce. The top cross is commonly used. A single self‐incompatible parent is used as female, and is open‐pollinated by a desirable cultivar as pollen source.
Male sterility
Male sterilityis a condition in plants whereby the anthers or pollen are nonfunctional. The condition may manifest most commonly as absence of/or extreme scarcity of pollen, severe malformation or absence of flowers or stamens, or failure of pollen to dehisce. Just like self‐incompatibility, male sterility enforces cross‐pollination. Similarly, it can be exploited as a tool to eliminate the need for emasculation for producing hybrid seed. There are three basic kinds of male sterility based on the origin of the abnormality:
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Principles of Plant Genetics and Breeding»
Представляем Вашему вниманию похожие книги на «Principles of Plant Genetics and Breeding» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Principles of Plant Genetics and Breeding» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.