Michael Begon - Ecology
Здесь есть возможность читать онлайн «Michael Begon - Ecology» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Ecology
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Ecology: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Ecology»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
– now in full colour – offers students and practitioners a review of the ecological sciences.
The previous editions of this book earned the authors the prestigious ‘Exceptional Life-time Achievement Award’ of the British Ecological Society – the aim for the fifth edition is not only to maintain standards but indeed to enhance its coverage of Ecology.
In the first edition, 34 years ago, it seemed acceptable for ecologists to hold a comfortable, objective, not to say aloof position, from which the ecological communities around us were simply material for which we sought a scientific understanding. Now, we must accept the immediacy of the many environmental problems that threaten us and the responsibility of ecologists to play their full part in addressing these problems. This fifth edition addresses this challenge, with several chapters devoted entirely to applied topics, and examples of how ecological principles have been applied to problems facing us highlighted throughout the remaining nineteen chapters.
Nonetheless, the authors remain wedded to the belief that environmental action can only ever be as sound as the ecological principles on which it is based. Hence, while trying harder than ever to help improve preparedness for addressing the environmental problems of the years ahead, the book remains, in its essence, an exposition of the
of ecology. This new edition incorporates the results from more than a thousand recent studies into a fully up-to-date text.
Written for students of ecology, researchers and practitioners, the fifth edition of
is anessential reference to all aspects of ecology and addresses environmental problems of the future.
Ecology — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Ecology», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
On the other hand, later in the main experiment ( Figure 5.28a) size inequality de creased in many cases. The reason is apparent in Figure 5.28c, which shows that the plants that died in the populations, ultimately, were the smallest ones. Thus, while size‐dependent growth‐differentials increased the size inequalities, size‐dependent survival‐differentials decreased them again by cutting off the tail of the size distribution. The modular nature of plants makes this separation of processes especially likely. There can be an extended period over which the smaller, weaker competitors are stunted in their growth but do not die, and only later, after the inequalities have been exaggerated, do differentials in survival reverse this. For most (unitary) animals, stunting is not an option and the weakest competitors die far sooner. As we have seen, however, patterns in animals like those seen in plants, while perhaps rarer, are certainly not unknown.
roots and shoots: the strength and asymmetry of competition
If competition is asymmetric because superior competitors pre‐empt resources, then competition is most likely to be asymmetric when it occurs for resources that are most liable to be pre‐empted. Specifically, competition amongst plants for light, in which a superior competitor can overtop and shade an inferior, might be expected to lend itself far more readily to pre‐emptive resource capture than competition for soil nutrients or water, where the roots of even a very inferior competitor will have more immediate access to at least some of the available resources than the roots of its superiors. This expectation is borne out by the results of an experiment in which morning glory vines ( Ipomoea tricolor ) were grown as single plants in pots (‘no competition’), as several plants rooted in their own pots but with their stems intertwined on a single stake (‘shoots competing’), as several plants rooted in the same pot, but with their stems growing up their own stakes (‘roots competing’) and as several plants rooted in the same pot with their stems intertwined on one stake (‘shoots and roots competing’) ( Figure 5.29). Root competition was more intense than shoot competition, in the sense that it led to a far greater decrease in the mean weight of individual plants. However, it was shoot competition for light that led to a much greater increase in size inequality and determined which individuals were dominant – a general conclusion drawn by Kiær et al . (2013) in their meta‐analysis of root and shoot competition.
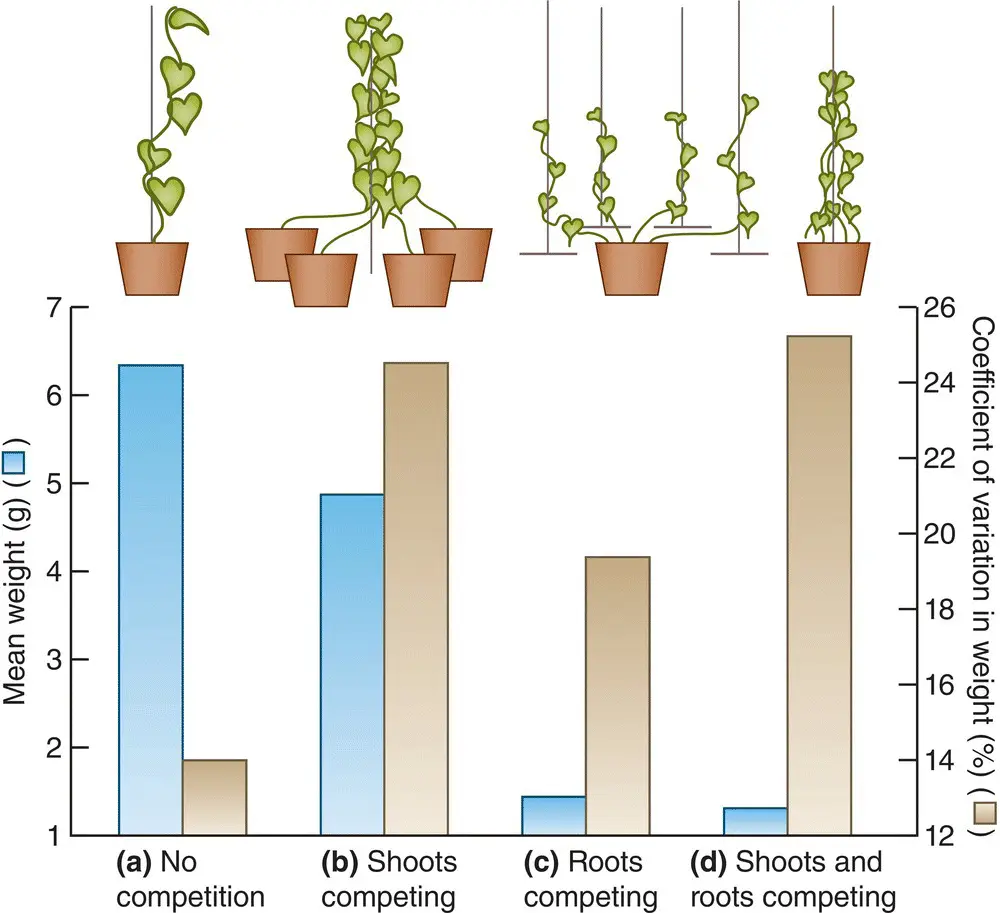
Figure 5.29 Root and shoot competition can have contrasting effects on mean size and size inequalities.When morning glory vines competed, root competition was most effective in reducing mean plant weight (treatments significantly different, P < 0.01, for all comparisons except (c) with (d)), but shoot competition was most effective in increasing the degree of size inequality, as measured by the coefficient of variation in weight (significant differences between treatments (a) and (b), P < 0.05, and (a) and (d), P < 0.01).
Source : After Weiner (1986).
5.8.3 Asymmetry enhances regulation
One final, important general point is that these asymmetries tend to reinforce the regulatory powers of intraspecific competition, in much the same way as exactly‐compensating or ‘contest’ competition ( Section 5.3) gives rise to the most tightly regulated populations. This can be seen especially in a much longer term study that will almost certainly never be repeated, where a population of the herbaceous perennial, Anemone hepatica , in Sweden was visited and revisited every year from 1943 to 1956, with each plant being tracked individually ( Figure 5.30; Tamm, 1956). Crops of seedlings entered the population each year between 1943 and 1952, but nonetheless, the most important factor determining which individuals survived to 1956 was whether or not they were established in 1943. Of the 30 individuals that were already of a large or intermediate size in 1943, 28 survived until 1956, and some of these had branched. By contrast, of the 112 plants that were either small in 1943 or appeared as seedlings subsequently, only 26 survived to 1956, and not one of these was sufficiently well established to have flowered. Tamm’s established plants were successful competitors – winners in a contest – year after year, but his small plants and seedlings were repeatedly unsuccessful. This guaranteed a near constancy in the number of established, winning plants between 1943 and 1956, accompanied by a variable number of ‘losers’ that not only failed to grow, but usually, in due course, died. Similar patterns can be observed in tree populations. The survival rates, the birth rates and thus the fitnesses of the few established adults are high; those of the many seedlings and saplings are low.
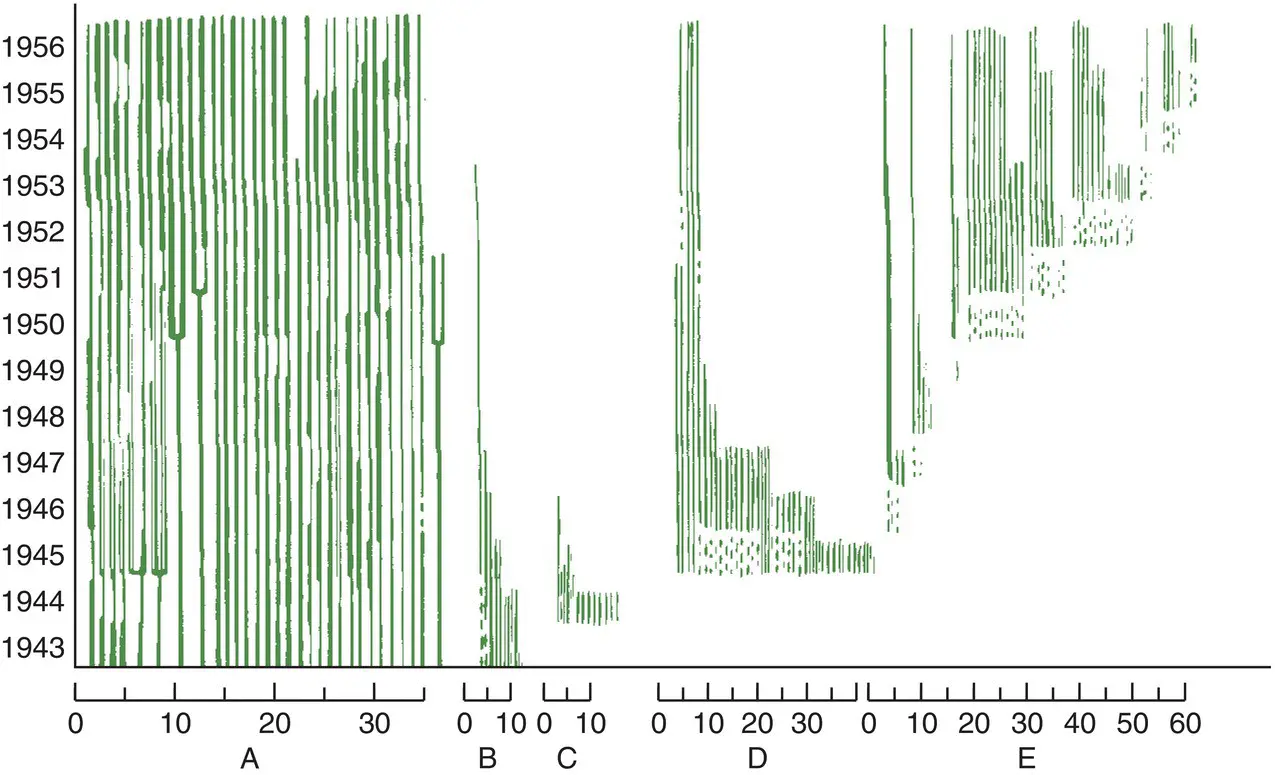
Figure 5.30 Asymmetric competition enhances population size regulation in an anemone.Space pre‐emption in a perennial, Anemone hepatica , in a Swedish forest. Each line represents one individual: straight for unramified ones, branched where the plant has ramified (i.e. produced new plants by vegetative growth), bold where the plant flowered and broken where the plant was not seen that year. Group A were alive and large in 1943, group B alive and small in 1943, group C appeared first in 1944, group D in 1945 and group E thereafter, presumably from seedlings.
Source : After Tamm (1956).
5.8.4 Territoriality
Territoriality refers to active interference between individuals, such that a more or less exclusive area, the territory, is defended against intruders by a recognizable pattern of behaviour. It is one widespread and important example of asymmetric intraspecific competition.
territoriality is a contest and regulates population size
Individuals of a territorial species that fail to obtain a territory, commonly referred to as ‘floaters’, often make no contribution whatsoever to future generations. Territoriality, therefore, is a ‘contest’. There are winners (those that come to hold a territory) and losers (those that do not), and at any one time there can be only a limited number of winners. The exact number of territories (winners) in a population may vary, depending on resources or environmental conditions. We see this, for example, for the number of tawny owl ( Strix aluco ) territories in a Finnish population, and especially the number of productive territories, which varies with the abundance of their vole prey ( Figure 5.31). Nonetheless, the contest nature of territoriality ensures, like asymmetric competition generally, a comparative constancy in the number of surviving, reproducing individuals. In the past, some felt that these regulatory consequences of territoriality must themselves be the root cause underlying the evolution of territorial behaviour – territoriality being favoured because the population as a whole benefitted from the rationing effects, which guaranteed that the population did not overexploit its resources (e.g. Wynne‐Edwards, 1962). However, there are powerful and fundamental reasons for rejecting this ‘group selectionist’ explanation (essentially, it stretches evolutionary theory beyond reasonable limits): the ultimate cause of territoriality must be sought within the realms of natural selection, in some advantage accruing to the individuals concerned.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Ecology»
Представляем Вашему вниманию похожие книги на «Ecology» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Ecology» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.