Michael Begon - Ecology
Здесь есть возможность читать онлайн «Michael Begon - Ecology» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Ecology
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Ecology: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Ecology»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
– now in full colour – offers students and practitioners a review of the ecological sciences.
The previous editions of this book earned the authors the prestigious ‘Exceptional Life-time Achievement Award’ of the British Ecological Society – the aim for the fifth edition is not only to maintain standards but indeed to enhance its coverage of Ecology.
In the first edition, 34 years ago, it seemed acceptable for ecologists to hold a comfortable, objective, not to say aloof position, from which the ecological communities around us were simply material for which we sought a scientific understanding. Now, we must accept the immediacy of the many environmental problems that threaten us and the responsibility of ecologists to play their full part in addressing these problems. This fifth edition addresses this challenge, with several chapters devoted entirely to applied topics, and examples of how ecological principles have been applied to problems facing us highlighted throughout the remaining nineteen chapters.
Nonetheless, the authors remain wedded to the belief that environmental action can only ever be as sound as the ecological principles on which it is based. Hence, while trying harder than ever to help improve preparedness for addressing the environmental problems of the years ahead, the book remains, in its essence, an exposition of the
of ecology. This new edition incorporates the results from more than a thousand recent studies into a fully up-to-date text.
Written for students of ecology, researchers and practitioners, the fifth edition of
is anessential reference to all aspects of ecology and addresses environmental problems of the future.
Ecology — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Ecology», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
3.8.2 Resource dimensions of the ecological niche
In Chapter 2we developed the concept of the ecological niche as an n ‐dimensional hypervolume. This defines the limits within which a given species can survive and reproduce for a number ( n ) of environmental factors, including both conditions and resources. Note, therefore, that the zero growth isoclines in Figure 3.30define niche boundaries in two dimensions. Resource combinations to one side of this zero isocline allow the organisms to thrive – but to the other side of the line the organisms decline.
The resource dimensions of a species’ niche can sometimes be represented in a manner similar to that adopted for conditions, with lower and upper limits within which a species can thrive. Thus, a predator may only be able to detect and handle prey between lower and upper limits of size. For other resources, such as mineral nutrients for plants, there may be a lower limit below which individuals cannot grow and reproduce but an upper limit may not exist ( Figure 3.30a–d). However, many resources must be viewed as discrete entities rather than continuous variables. Larvae of butterflies in the genus Heliconius require Passiflora leaves to eat; those of the monarch butterfly specialise on plants in the milkweed family; and various species of animals require nest sites with particular specifications. These resource requirements cannot be arranged along a continuous graph axis labelled, for example, ‘food plant species’. Instead, the food plant or nest‐site dimension of their niches needs to be defined simply by a restricted list of the appropriate resources.
Together, then, conditions and resources define a species’ niche. In the next chapter, we will look in more detail at the most fundamental responses of organisms to those conditions and resources: their patterns of growth, survival and reproduction.
3.9 A metabolic theory of ecology
Resources and conditions are also important insofar as they determine the metabolic rates of individuals, which determine the levels of resources available to those individuals for reproduction, growth and so on, which in turn influence their life histories, their abundance and indeed all of the processes we discuss in subsequent chapters. This perspective has generated interest in a ‘metabolic theory of ecology’ (Brown et al ., 2004). At the heart of metabolic theory are the effects on metabolic rate of temperature, and in particular of size (most often, body mass). We examined the effects of temperature in Section 2.3. We turn now to size.
metabolic scaling
The most fundamental point, perhaps, is that life is typically faster for small organisms than it is for large ones – metabolising at greater rates, and maturing and dying sooner. For example, per gram of body mass, a resting mouse metabolises about 20 times faster than an elephant. There are exceptions to this pattern, as we’ll see below, but the more general rule is very widespread. Putting this more formally, we can say that the rate of a metabolic process, Y , varies with size according to the equation
(3.1) 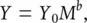
where Y 0is referred to as the normalisation constant, reflecting the typical rate for the metabolic process concerned, M is the organism’s mass and b is the so‐called allometric exponent. Equation 3.1may be said to describe metabolic scaling . Taking logs of both sides, we get:
(3.2) 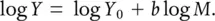
allometric relationships
The slowing down of metabolism at larger sizes is reflected in values of b that are less than 1. This makes them allometric relationships – that is, relationships in which a physical or (in this case) physiological property of an organism changes relative to the size of the organism, rather than changing in direct proportion to the changing size. That would be an iso metric relationship, and in that case b would be equal to 1. We can see from Equation 3.2that b is the slope of the line when metabolism is plotted against body mass on logarithmic scales.
Examples are shown in Figure 3.31for a wide range of taxonomic groups. As mass increases, temperature‐corrected metabolic rate increases less than proportionately; b = 0.71, with, apparently, little variation between groups. We discuss actual values of b below. The intercept in these plots is log Y 0, such that the value of Y 0locates the relationship vertically within the plot, telling us about the absolute rate of metabolism at a given body size (and temperature). In this case, there is variation in absolute rate between the groups, despite the relationship with size being apparently the same. Such allometric relationships can be ontogenetic (changes occurring as an organism grows) or phylogenetic (changes that are apparent when related taxa of different sizes are compared).
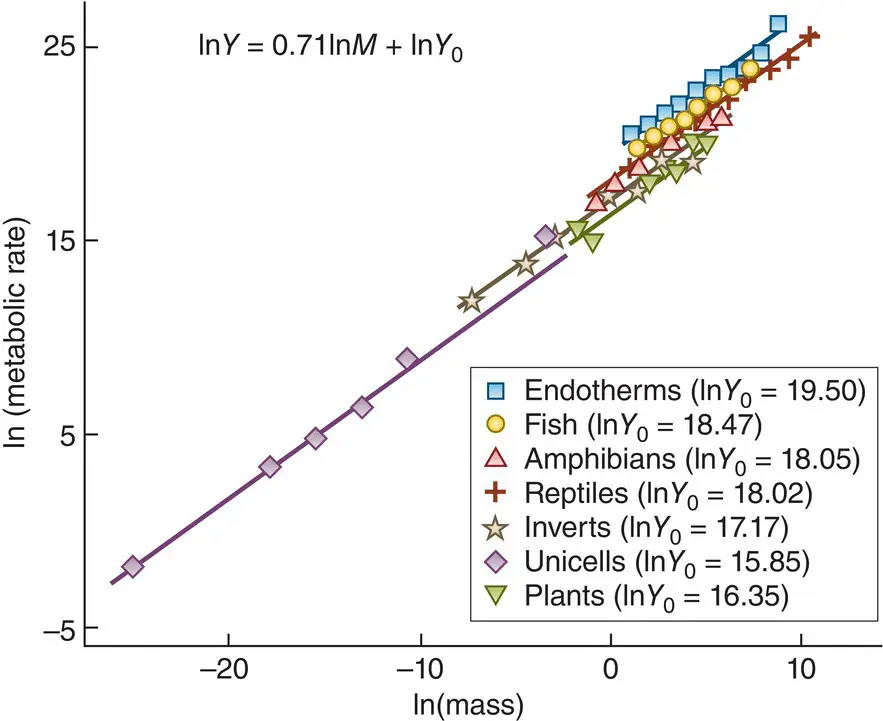
Figure 3.31 Metabolic scaling: the relationship between metabolic rate (Y, watts) and body mass (M, g) for a variety of organisms, as indicated, on logarithmic scales.The analysis sought a single slope but allowed the intercepts of different groups to vary. For clarity, the data points shown are the averages for mass‐classes. The metabolic rates have been temperature‐corrected to ensure different studies are comparable (see Equation 3.3), but this has been omitted from the equation, top right, for clarity.
Source : After Brown et al . (2004).
rates per unit mass, and times
Note that if individual metabolic rate scales with individual mass with an exponent of b , then metabolic rate per unit mass (that is, the metabolic rate of a gram of tissue) will scale with an exponent of b –1 (simply divide both sides of Equation 3.1by M ). Similarly, the time taken to complete a process (for example to reach maturity) will scale with an exponent of 1– b , because these times are the reciprocals of rates per unit mass (the reciprocal exponent of b –1 is –( b –1) = 1– b ). It may sometimes be more appropriate to examine the metabolic scaling of times or rates per unit mass than individual rates.
transport or demand?
The conventional view, as we discuss later, is that plots like Figure 3.31arise because size imposes constraints on rates of supply (of oxygen, nutrients and so on), and of transport of materials generally within the organism, which determine metabolic rates. Metabolic rates then constrain the resources available for reproduction, growth and so on (Brown et al ., 2004). An alternative viewpoint, however, is that size is closely co‐adapted with investment in reproduction and growth (and hence with the demands for these), with rates and means of transport evolving to satisfy those demands (Harrison, 2017). Size in this case is an integral part of an organism’s overall life history, which has evolved to match its environment. From this perspective, a mouse, for example, metabolises rapidly, and an elephant metabolises more slowly, to fuel their respective life histories, of which size is a part. Each has evolved transport networks sufficient to service their metabolism. We discuss life history evolution more fully (and why mice might be fast, and elephants slow) in Chapter 7. The alternative viewpoints are summarised in Figure 3.32. Bringing the two together, we may conclude that there is really no single driver of metabolic scaling. Rather, we should see life histories and their metabolic demands as being co‐adjusted with transport system design (Glazier, 2014).
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Ecology»
Представляем Вашему вниманию похожие книги на «Ecology» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Ecology» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.