Philias R. Garant - Oral Cells and Tissues
Здесь есть возможность читать онлайн «Philias R. Garant - Oral Cells and Tissues» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Oral Cells and Tissues
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Oral Cells and Tissues: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Oral Cells and Tissues»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Oral Cells and Tissues — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Oral Cells and Tissues», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
226. Veis A. Phosphoproteins of dentin and bone: Do they have a role in matrix mineralization? In: Butler WT (ed). The Chemistry and Biology of Mineralized Tissues. Birmingham, AL: EBSCO Media, 1985:170–176.

Enamel
Differentiation of the Enamel Organ
During the early stage of tooth formation, the enamel organ consists of the outer enamel epithelium (OEE), the cells of the stellate reticulum (SR), the stratum intermedium (SI), and the inner enamel epithelium (IEE) (see Fig 1-6). The cells of the outer enamel epithelium are generally cuboidal. They attach by hemidesmosomes to a basal lamina separating them from the adjacent dental sac, a connective tissue of ectomesenchymal origin. Cytoplasmic organelles in the OEE include a moderate number of mitochondria, a small number of cisterns of rough endoplasmic reticulum (RER) and smooth endoplasmic reticulum, and a poorly developed Golgi complex. The presence of coated vesicles in the peripheral cytoplasm and along the plasma membrane facing the basal lamina suggests that the OEE is involved in specific endocytosis of extracelluar substances.
Soon after the onset of enamel formation, the OEE becomes convoluted by indentations of highly vascularized connective tissue. This structural change becomes pronounced during enamel maturation, when the OEE, the SR, and the SI form the papillary layer to increase the surface area between the enamel organ and the adjacent blood supply. This change is pronounced in the continuously developing incisor of the rat, the most thoroughly investigated model of tooth development.
The cells of the SR have a compact cell body with many long folds of cytoplasm that contact and partly overlap similar cytoplasmic folds from adjacent cells. Desmosomes and gap junctions are formed at points of cell-to-cell contact. These folds of cytoplasm line wide extracellular spaces rich in water-binding glycosaminoglycans.
The soft, jellylike consistency of the enamel organ is due to the hydration of the SR glycosaminoglycans. This property is believed to be important in equalizing pressure generated by cell proliferation and matrix secretion in the dental papilla. It has been suggested that, if external tissue-generated forces at the interface between the preameloblasts and the preodontoblasts are eliminated, the three-dimensional outline of the future dentinoenamel junction, and ultimately the shape of the crown of the tooth, can be molded solely by the forces of cell proliferation in both the IEE and the underlying preodontoblasts of the dental papilla. With the onset of ameloblast differentiation, the formation of a terminal web in the IEE, coupled with the assembly of a basement membrane beneath the IEE, stabilizes the shape of the future dentinoenamel junction.
The cells of the SI form a compact zone, one to two cellular layers deep, between the SR and the IEE ( Figs 3-1and 3-2). The intercellular spaces between the SI cells are relatively narrow, and the adjacent cells are held together by many desmosomes. Large gap junctions are formed with adjacent cells of the IEE. The SI cells contain many mitochondria, a characteristic shared with the distal cytoplasm of secretory ameloblasts.
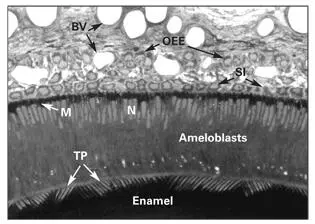
Fig 3-1Secretory ameloblasts of the rat incisor tooth. In the secretory stage, ameloblasts are tall columnar cells, characterized by a secretory process (Tomes process [TP]) and an infranuclear concentration of mitochondria (M). (BV) Blood vessels; (N) nucleus; (OEE) outer enamel epithelium; (SI) stratum intermedium.
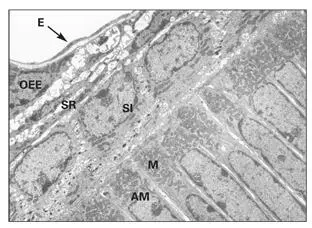
Fig 3-2Electron micrograph of the proximal end of secretory ameloblasts (AM) containing a high concentration of mitochondria (M). Cells of the stratum intermedium (SI), stellate reticulum (SR), and outer enamel epithelium (OEE) have not yet formed the papillary layer. Blood vessels (endothelium [E]) lie in close apposition to the OEE. (Original magnification × 1,600.)
Although the exact function of the cells of the SI is not clear, the fact that they contain a rich complement of mitochondria and stain intensely for alkaline phosphatase and adenosine triphosphatase (ATPase), suggests a possible role in ion and water transport. The presence of extensive gap junctions between the cells of the SI and the ameloblasts indicates that the two cell types act in close concert during amelogenesis.
Recent studies have shown that the regulatory activity of the enamel knot is taken up by the SI during cusp formation. 1Growth regulatory signals mediated by Sonic Hedgehog (Shh) are activated in the SI in a wavelike manner from the cusp to cervical zone.
The IEE is a layer of columnar preameloblasts abutting the dental papilla. Organ culture studies of amelogenesis indicate that contact with the dental papilla is required for the expression of enamel protein. 2Although the precise nature of the early instructive signals that originate from the dental papilla have yet to be identified, the permissive effect of the extracellular matrix of the basement membrane is required for initiating expression of amelogenin. 3
The region of highest mitotic activity in the IEE is located near the cervical loop portion of the enamel organ. 4– 6Stem cells in the cervical loop divide to give rise to an expanding metablast clone of preameloblasts. 7, 8The rates of proliferation and differentiation in the blast-metablast populations vary among species, among individual teeth, and among different parts of a given tooth.
Secretion of growth factors by the enamel knot and the dental papilla regulate cell proliferation and growth of the IEE. 6, 9Receptors for epidermal growth factor, platelet-derived growth factor, and fibroblast growth factor have been localized by immunohistochemistry in preameloblasts. 10Phospholipase C γ, a downstream signaling molecule activated by growth factor–receptor interaction, was also demonstrated in preameloblasts. Preameloblasts must exit the cell cycle to begin the process of cytodifferentiation. 4
The cytoplasm of the preameloblasts contains many free ribosomes, a small Golgi apparatus, a few cisterns of rough and smooth endoplasmic reticulum, and a small number of mitochondria. Adjacent preameloblasts form gap junctions and desmosomes. A prominent zonula adherens junction with an associated terminal web of cytoplasmic filaments forms in the proximal cytoplasm adjacent to the cells of the SI. 11, 12This proximal junctional complex binds and stabilizes the preameloblasts prior to degradation of the underlying basal lamina.
Soon after the deposition of mantle dentin, the odontoblasts express matrix metalloproteinases (MMPs) that begin the digestion of the basement membrane. Preameloblasts subsequently remove basement membrane fragments through phagocytosis. 13Removal of the basement membrane allows the dentin and enamel matrices and their respective mineral phases to come into direct contact. As a result, a strong mechanical bond is formed between the enamel and dentin.
Recent studies using reverse transcription and polymerase chain reaction techniques have shown that immature enamel organ cells exist in a “protodifferentiated” state. 14Preameloblasts of the IEE produce small amounts of enamel matrix protein prior to overt morphodifferentiation as secretory phenotypes. 15, 16Some of these proteins traverse the dental lamina and are taken up in odontoblast-coated vesicles.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Oral Cells and Tissues»
Представляем Вашему вниманию похожие книги на «Oral Cells and Tissues» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Oral Cells and Tissues» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.