Julio C. Rojas - Temas selectos en ecología química de insectos
Здесь есть возможность читать онлайн «Julio C. Rojas - Temas selectos en ecología química de insectos» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на испанском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Temas selectos en ecología química de insectos
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Temas selectos en ecología química de insectos: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Temas selectos en ecología química de insectos»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Temas selectos en ecología química de insectos — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Temas selectos en ecología química de insectos», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Los machos de las diferentes familias de lepidópteros emplean estrategias específicas para localizar las fuentes de feromona. Sin embargo, todas las especies estudiadas despliegan un comportamiento de orientación característico hacia las fuentes de feromona basado en anemotaxias optomotoras (Kennedy & Marsh, 1974). Es decir, emplean imágenes visuales en movimiento para dirigir una trayectoria, con una velocidad particular con respecto al viento (Baker, 1986; Arbas, 1997; Baker & Vickers, 1997; Cardé & Mafra-Neto, 1997; Willis & Arbas, 1997; Witzgall, 1997).
Una vez que son expuestos a las feromonas los machos de lepidópteros, y también los de muchos otros ordenes de insectos, emprenden el vuelo en dirección contraria al viento en un patrón ondulante con trayectorias cortas y pequeños ángulos con respecto a la dirección principal de la estela de olor. Si pierden contacto con la estela presentan patrones de locomoción con giros largos y ángulos grandes que cruzan la trayectoria principal de la estela, con poco o ningún desplazamiento, hasta establecer nuevamente contacto con la estela (Kennedy, 1983) (Figura 2).

Figura 2.Vuelo en dirección contraria al viento en un patrón ondulante con trayectorias cortas y pequeños ángulos con respecto a la dirección principal de la estela de olor.
El comportamiento mostrado es específico para cada especie y depende de la concentración de feromona y la forma en la que ésta es emitida (Cardé & Hagaman, 1979; Kuenen & Baker, 1982; Willis & Arbas, 1991; Justus & Cardé, 2002). Se ha propuesto la existencia de un mecanismo generador de las trayectorias de vuelo, que es independiente del viento y disparado por estímulos intermitentes de olor (Kennedy et al ., 1980; Baker & Kuenen, 1982; Kuenen & Baker, 1983; Baker et al ., 1984; Vickers, 2000; Vickers, 2006; Cardé & Willis, 2008). El patrón de vuelo que resulta de este generador de vuelo consiste principalmente de dos elementos básicos: i) trayectorias cortas en zig-zag con ángulos pequeños y ii) trayectorias relativamente más largas y con ángulos mayores, ambas generadas internamente inmediatamente después del primer contacto con el estímulo de la feromona. La intermitencia de la estimulación parece ser necesaria para mantener la trayectoria en el movimiento de orientación hacia la fuente de feromona. De hecho, la localización eficiente de la fuente de feromona depende de la concentración, temporalmente fluctuante, de la misma feromona encontrada por una palomilla (Kennedy et al. , 1980; Willis & Baker, 1984; Baker et al. , 1985). El número de giros (cambios de dirección) disminuye conforme incrementa la frecuencia del pulso de olor, dando lugar a trayectorias rectas de vuelo que permiten a la palomilla alcanzar la fuente de olor (Mafra-Neto & Cardé 1994; Vickers & Baker 1994) (Figura 3).
Finalmente, se ha propuesto que las trayectorias largas son iniciadas por estelas filamentosas de la feromona y las trayectorias cortas son iniciadas por la presencia de bolsas de aire limpio, de suficiente duración, en la estela principal (Baker, 1990). Es necesario mencionar que algunas palomillas muestran giros espontáneamente sin un estímulo olfatorio contínuo y su comportamiento sugiere que la localización de la fuente de olor se logra por un patrón activamente generado de giros y contragiros, continuamente modulado por la información de diferentes modalidades sensoriales que recibe el insecto (Kaissling & Kramer, 1990; Willis & Arbas, 1997, 1998).
La anemotaxia optomotora, la combinación de los tipos de trayectoria generadas internamente, así como el patrón temporal de estimulación de la feromona, se considera que dan lugar al comportamiento de vuelo observado en este grupo de insectos. Sin embargo, algunos aspectos de este esquema, particularmente la existencia de un mecanismo generador de giros y contra-giros, siguen en discusión (Arbas, 1997; Baker & Vickers, 1997; Cardé & Mafra-Neto, 1997). Estos aspectos fueron recientemente revisados por Cardé & Willis (2008) (Figura 3).
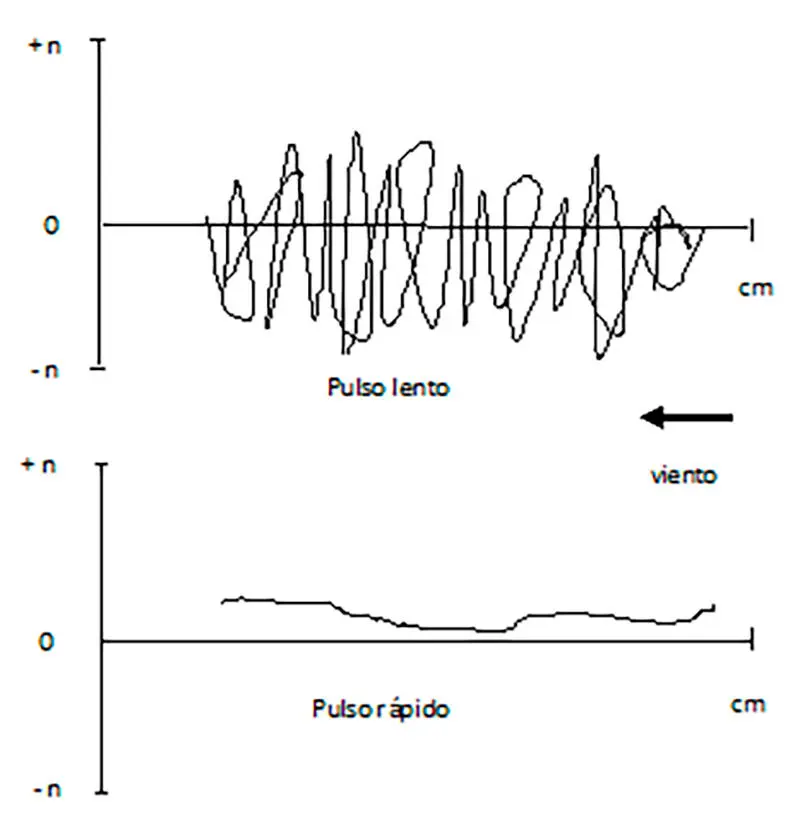
Figura 3.Tipos de vuelo presentados ante pulsos lentos y pulsos rápidos de olor. El número de giros (cambios de dirección) disminuye conforme incrementa la frecuencia del pulso de olor, dando lugar a trayectorias rectas de vuelo.
Después de que un macho localiza una hembra, varias señales, incluyendo a las feromonas, están involucradas en el comportamiento que precede a la cópula (Krasnoff et al ., 1987; Charlton & Cardé, 1989). El macho no volador de B. mori ha sido particularmente útil para investigar la orientación a la feromona en individuos no voladores en donde los movimientos son restringidos a dos dimensiones (Kanzaki, 1997). A pesar de las similitudes con las palomillas voladoras, los mecanismos no son necesariamente idénticos (Kramer, 1997). Por ejemplo, en respuesta a la feromona los machos de Grapholita molesta muestran giros y contragiros mientras se orientan en el viento y no directamente al caminar (Willis & Baker, 1987). El patrón de locomoción de B. mori en la orientación respecto a la feromona depende de la estructura temporal de la estimulación (Kramer, 1975, 1986, 1992; Kanzaki et al ., 1992); con una estimulación con pulsos de olor, como la que ocurre en condiciones naturales, se observa un patrón de marcha en zig-zag contra el viento, lo cual también es apoyado por un desplazamiento inducido por el batir de las alas, el cual se presenta incluso después de la ablación de las patas (Kanzaki, 1998). Como en las palomillas voladoras, en esta especie hay evidencia que sugiere la existencia de un generador interno de giros (Kanzaki et al ., 1992).
La secuencia básica del comportamiento inducido por pulsos de feromona, consiste de una trayectoria larga recta seguida por giros y contragiros, y finalmente por vueltas completas. A partir de un cierto intervalo de frecuencia de estimulación se induce una marcha en zig- zag cuya trayectoria se convierte más recta conforme aumenta la frecuencia de estimulación. El patrón de locomoción está asociado con las posturas de antenas, abdomen y alas (Olberg, 1983; Kanzaki, 1998; Mishima & Kanzaki, 1998).
Respuestas de las hembras a feromonas de machos
Las feromonas de los machos estimulan en las hembras comportamientos de búsqueda y localización de pareja, tales como marcha dirigida y atracción a corta distancia, y aceptación y receptividad (Birch et al. , 1990). La confirmación experimental de estas funciones, sin embargo, ha sido más difícil que en el caso de las feromonas producidas por hembras, debido principalmente a que las feromonas de machos no tienen una atracción a larga distancia y las determinaciones de su estructura han sido reportadas sin proporcionar detalles acerca de su actividad biológica. En el caso de E. excrescens , observaciones del comportamiento de apareamiento, realizadas en campo, confirmaron la existencia de un vuelo dirigido de tipo pendular hacia el macho por parte de la hembra e indicaron que la hembra fue atraída por el macho por estímulos visuales a grandes distancias y por la feromona secretada a corta distancia (Kan et al ., 2002).
Plasticidad en las respuestas a feromonas
Las respuestas a las feromonas aunque esterotipadas y especie-específicas no son rígidas. En diferentes condiciones pueden presentar variaciones en la sensibilidad comportamental del individuo receptor. Estas variaciones en la sensibilidad a las feromonas pueden ocurrir durante la interacción de éstas con otros olores diferentes (e.g. volátiles de plantas) o la interacción de las feromonas con otras modalidades sensoriales (e.g. colores, formas) (Landolt et al ., 1994; Rojas & Wyatt, 1999). Adicionalmente la existencia de ritmos endógenos pueden afectar las respuestas de locomoción guiadas por las feromonas (Silvegren et al ., 2005). Asimismo el aprendizaje puede jugar un papel importante en la generación de variación en la respuesta a las feromona. Aprendizaje de tipo no asociativo puede presentarse debido a la repetición en la exposición al olor de la feromona, dependiendo de la madurez o estado fisiológico del insecto (Judd et al ., 2005) y en algunos casos también procesos de aprendizaje asociativo pueden afectar las respuestas a las feromonas (Skiri et al ., 2005).
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Temas selectos en ecología química de insectos»
Представляем Вашему вниманию похожие книги на «Temas selectos en ecología química de insectos» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Temas selectos en ecología química de insectos» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.