Joseph J. Torres - Life in the Open Ocean
Здесь есть возможность читать онлайн «Joseph J. Torres - Life in the Open Ocean» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Life in the Open Ocean
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Life in the Open Ocean: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Life in the Open Ocean»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Life in the Open Ocean: The Biology of Pelagic Species
Life in the Open Ocean: The Biology of Pelagic Species
Life in the Open Ocean — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Life in the Open Ocean», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Narcomedusae
Solmundella bitentaculata ( Figure 3.17d) has two tentacles that are held in front of the bell (aboral side) as it swims, forming a cylindrical or conical volume of about three bell diameters in front of the bell.
Aeginopsis laurentii is another D‐type ( Figure 3.17d) predator but with four tentacles that extend out in front of the bell in a cone. Like Solmundella , Aeginopsis is a mesopelagic species and little information is available on diet.
Solmaris is a narcomedusan genus with several species living at epi‐ and mesopelagic depths. It conforms to the narcomedusan D‐type tentacle deployment; it has an array of several tentacles forming a larger cone.
Scyphomedusae
Pelagia noctiluca is a semeaostome scyphomedusa. As a scyphozoan, it is larger than its hydrozomedusan brethren with a bell diameter of up to 15 cm and tentacles extending outward up 30 times the bell diameter. It swims continuously as a C‐type predator feeding on a wide variety of prey types including salps, doliolids, pteropods, forams, amphipods, chaetognaths, and fishes.
The Model
Madin (1988) proposed a general model to describe tentaculate predation incorporating the best elements of previous studies (Gerritsen and Strickler 1977; Mills 1981; Greene 1985; Greene et al. 1986; Larson 1987a, 1987b; Purcell and Mills 1988) and the author’s own observations. A successful predatory sequence consists of four parts: encounter, capture/selection, ingestion, and digestion.
Interaction with prey begins with the “encounter” phase, which is determined by the size and type of encounter zone ( Figure 3.17a–d) and the swimming behavior of the medusa. It was assumed that sensory mechanisms were not used to target individual prey. Once a prey item is within its encounter zone, how effective a species will be at capturing it will be determined by a suite of characteristics: tentacle density and spacing, prey size and swimming behavior, and effectiveness of the nematocysts and venom.
Prey size is dictated to some degree by the morphology of the predator. Species with fine dense tentacles (A‐type in Figure 3.17) may be expected to capture small prey items, e.g. copepods, and those with more widely spaced robust tentacles (B and C‐type) to capture larger prey, e.g. ctenophores and other medusae. Many of the B‐ and C‐type predators increase their encounter probability with prey using swimming or hunting behavior. The effectiveness of the nematocysts in paralyzing prey and strength of the tentacles for retaining larger prey are also important in the successful capture of larger prey items. Species that prey on soft‐bodied species such as jellies and fishes have one type of nematocyst designed to harpoon and poison the prey, whereas those that prey on crustaceans have four to five types of nematocysts in specialized batteries designed to entangle and hold crustaceans. Clearly, those species (e.g. cubomedusae) that prey on strong swimmers such as fishes and larger shrimp that could potentially tear tentacles must have venom virulent enough to quickly paralyze their prey.
Once a prey item is captured, successful digestion will depend on whether the item can be conveyed to the mouth by the tentacles and whether it can be successfully introduced to the gastric cavity through the mouth and broken down into useful nutrients by the digestive apparatus.
Swimming and Hunting Behavior
Mills (1981) gives an example of four hydromedusan hunting behaviors ( Figure 3.18) that correspond well with Madin’s general model. The first species, Proboscidactyla flavicirrata ( Figure 3.18a) corresponds to the A‐type predator in Figure 3.18: neutrally buoyant with 40–80 tentacles radiating from a globular bell. P. flavicirrata employs a motionless ambush strategy, allowing small zooplankton to enter the “encounter zone” through their own swimming behavior. Its neutral buoyancy makes it an especially effective trap for its plankton prey.
As a congener of Stomotoca pterophylla , Stomotoca atra is a B‐type (see Figure 3.18b) predator. By adding its swimming behavior, we gain a better understanding of how it uses its two long tentacles to hunt. It employs a hop–sink swim cycle to drag its long tentacles up and down through the water column. As it swims and sinks, the tentacles describe a sine curve about the width of the bell and 2 meters from top to bottom. It feeds on large prey (hydromedusae), and it greatly increases the probability of contacting a prey item by its method of interrogating the water column.
The third species, Phialidium gregarium , is the primary prey of Stomotoca atra . P. gregarium also employs a hop–sink feeding strategy but a very different one from that of S. atra . It swims upward, bell uppermost, then sinks down with the bell oriented downward and its tentacles trailing behind ( Figure 3.18c). As it does so, vortices are created behind the bell that circulate small prey into the tentacles. P. gregarium would be classified as a C‐type predator in Madin’s Figure 3.17model.
The last example is Polyorchis pencillatus , a resident of shallow bays where it spends a great deal of its time on the bottom. Its hunting strategy on the bottom is to perch on its tentacles ( Figure 3.18e) and use its manubrium to ingest prey from the surface sediments. At intervals it hops up off the bottom, stirring up the sediments, and then back down. Occasionally it swims up to the surface and drifts downward bell up ( Figure 3.18d), becoming an A‐type predator in the Figure 3.17model.
Mills (1981) lists seven factors that contribute to feeding efficiency in medusae: (i) tentacle number and length; (ii) geometry of tentacle posture; (iii) velocity of tentacles moving through water; (iv) swimming pattern of medusa; (v) streamlining effects of the medusa bell on water flow; (vi) diameter of the prey; (vii) swimming pattern and velocity of prey. Together, Mill’s observations and Madin’s conceptual treatment provide a useful framework for examining the feeding strategies of medusae.
Water Flow and Swimming
The specialized swimming behaviors described above for the hydromedusae increase their hunting efficiency considerably. However, even conventional “straight‐line” swimming creates flow fields around the swimming medusa that enhance prey capture. Analysis of movement in the disc‐like moon jelly Aurelia aurita shows that normal swimming motions, particularly the expansion of the swimming bell during the recovery stroke, entrain particles to within easy reach of the tentacles as the water rushes into the subumbrellar space (cf. Costello 1992).
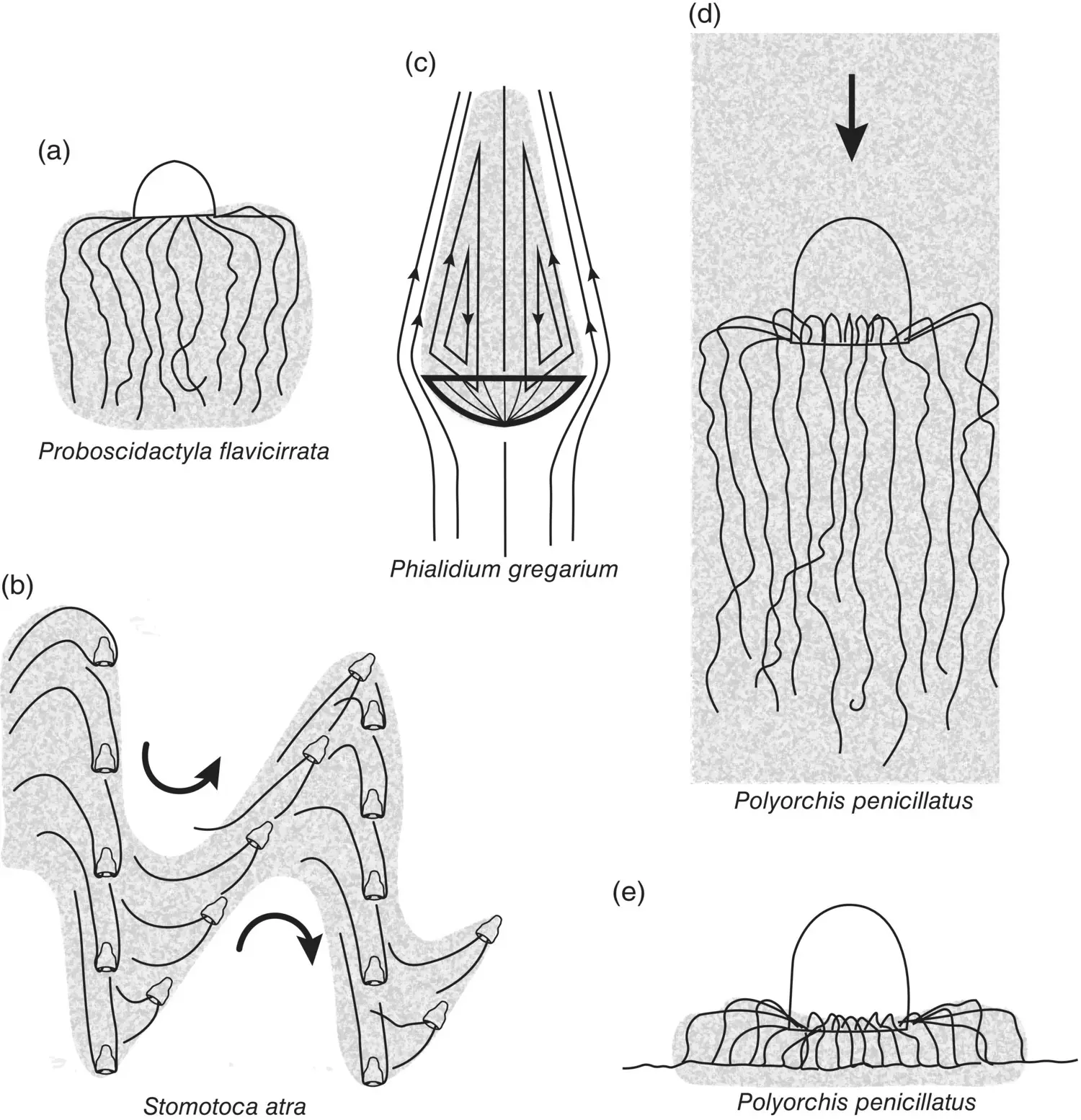
Figure 3.18 Different hunting and feeding behaviors of medusae. Shaded areas indicate effective feeding spaces. (a) Motionless ambush strategy; (b) swimming‐sinking search pattern; (c) downward sinking with trailing tentacles creating vortices, which bring food particles within the feeding space; (d and e) two strategies employed by medusae such as Polyorchis penicillatus; part‐time swimming and fishing in the water column and part‐time resting on its tentacles on the bottom and capturing the prey directly with its manubrium.
Source: Adapted from Mills (1981).
Attraction Between Predator and Prey
Attraction between predators and their prey can take several forms. Siphonophores create “lures” with nematocyst batteries on the fishing tentacles to attract the prey (Purcell 1980). In a different form of attraction, medusae have been observed to swim toward areas of high prey concentration (Arai 1991) in the laboratory, as well as aggregating in areas where prey have recently been, suggesting a chemoreceptive mechanism of attraction resulting in higher densities of medusae.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Life in the Open Ocean»
Представляем Вашему вниманию похожие книги на «Life in the Open Ocean» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Life in the Open Ocean» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.