Alan Gunn - Parasitology
Здесь есть возможность читать онлайн «Alan Gunn - Parasitology» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Parasitology
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Parasitology: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Parasitology»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Highly detailed textbook on parasites and parasite relationships Parasitology: An Integrated Approach
Parasitology: An Integrated Approach, 2nd edition
Parasitology — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Parasitology», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
4.2.1.3 Post Kala‐Azar Dermal Leishmaniasis
Post kala‐azar dermal leishmaniasis (PKDL) is a condition that is usually associated with L. donovani and develops as sequel to visceral leishmaniasis in 2.5–20% of cases (hence ‘post‐kala‐azar’). It may manifest itself anything from immediately afterwards to several years following the condition. It is characterised by the development of nodules and/or macules that can be extensive and cover any area of the body and may be mistaken for leprosy. The nodules are irregular raised masses on the skin surface whilst macules (Latin macula = blemish or small spot) are flat discoloured areas on the skin surface. These regions contain numerous amastigotes, and if they occur on exposed parts of the body, they are a ready source of infection for sandflies. The identification and treatment of patients suffering from PKDL is therefore an important part of any control programme. There are marked differences the occurrence and development of PKDL between countries, which suggests that host, and/or parasite factors may be important in whether it develops. For example, most cases of PKDL (~50%) occur in the Sudan and the condition tends to develop more rapidly there than in India.
Some workers consider that the development of PKDL is associated with the incomplete or inefficient treatment of visceral leishmaniasis following treatment with the drugs sodium stibogluconate and pentamidine. However, PKDL may also occur after treatment with miltefosine. In India, Das et al. (2009) found about 20% of PKDL cases occurred in people for whom there was no record of either visceral leishmaniasis or the prescription of the drugs used to treat it. However, these people tested positive for L. donovani and therefore carried an asymptomatic infection.
4.2.1.4 Cutaneous Leishmaniasis
Cutaneous leishmaniasis manifests itself in a variety of different forms depending upon the species of Leishmania . Clinically, cutaneous leishmaniasis divides into three basic types depending upon how the disease presents. Localized Cutaneous Leishmaniasis (LCL) generally takes the form of a dry ulcer. It develops at the bite site of the sandfly vector and usually heals by itself although this may take some time and leave permanent scarring. This form of disease is common in India, Central Asia, the Middle East, and parts of southern Europe; L. major and L. tropica are responsible for most infections in these regions. LCL is also common in South America where species such as L. venezuelensis and L. mexicana are responsible. Diffuse (disseminated) cutaneous Leishmaniasis (DCL) is a rarer and much more serious condition than LCL. It manifests as numerous raised (but not ulcerating) papules and nodules that spread to cover large areas of the body. The condition is often associated with immune suppression, and there are several reports of HIV co‐infection (e.g., Corrêa Soares et al. 2020). Unlike LCL, patients with DCL seldom recover without treatment. In the ‘Old World’, Leishmania aethiopica is the most common cause of DCL whilst in South America L. mexicana and L. amazonensis are implicated. As mentioned earlier, any one species of Leishmania may cause different types of leishmaniasis.
Mucocutaneous leishmaniasis (MCL) arises from the formation of an ulcerative lesion that afflicts the mouth, palate, and nose. As a rule, MCL develops and spreads slowly over a period of years and eventually destroys the affected region. It is most common in South America, particularly Brazil and the Amazon regions of Peru, Ecuador, Colombia, and Argentina where the condition is known as ‘espundia’. The discovery of pre‐Inca pottery illustrating disfigured faces suggests that the disease pre‐dates the European invasion of South America. However, some workers claim that the conquistadors and early Spanish settlers introduced MCL into South America. Espundia has a low mortality (~5%) provided the patient receives medical care. However, in the absence of treatment, mortality but would be undoubtedly higher. Death from MCL commonly results from complications such as aspiration pneumonia although some sufferers suffocate owing to laryngeal closure. Leishmania braziliensis is responsible for most cases of MCL although L. guyensis is also important. Leishmania major and some other species of Leishmania can also cause MCL, but these cases are rare. Leishmania guyanensis and L. braziliensis are naturally infected with Leishmania RNA virus 1 (Cantanhêde et al. (2018), and this results in the development of a particularly rapid and aggressive form of MCL (Olivier and Zamboni 2020).
4.2.2 Genus Trypanosoma
All members of this genus are parasites of vertebrate animals, and they are almost all transmitted by invertebrate vectors. Asexual reproduction usually takes place in both the vertebrate host and the invertebrate vector (i.e., the parasites are heteroxenous). However, in mechanically transmitted trypanosomes, such as Trypanosoma evansi , there is no development outside the vertebrate host. There is evidence of sexual reproduction (or something like it) occurring in the invertebrate host in some trypanosome species. Trypanosoma equiperdum is something of an exception to this general lifestyle because it is a sexually transmitted parasite of horses and other equine species. Nevertheless, most of the genus Trypanosoma are heteroxenous and two distinct groups are identifiable based on where they develop within their invertebrate host. Those developing within the anterior regions of the gut and transmitted when the vector bites exhibit ‘anterior station’ development and belong to the ‘Salivaria’ group. Species developing in the vector’s hindgut and transmitted via its faeces exhibit ‘posterior station development’ and belong to the ‘Stercoraria’ group. Both groups include species of medical and veterinary importance ( Table 4.4; Figure 4.6). It should come as no surprise that the taxonomy of trypanosomes undergoes regular rearrangements. It is highly likely that some well‐known species are actually ‘sub‐species’ or ‘synonyms’, but for the sake of comparison with past literature we have retained the most common species’ names.
In several trypanosome species, the kinetoplast is modified or lacking entirely. This has had a dramatic effect on their subsequent spread and evolution. In the case of Trypanosoma brucei , the development of the mitochondrion undergoes down regulation within the vertebrate host. However, the mitochondrion is fully functional during the procyclic stage that develops in the tsetse fly vector ( Figure 4.7). The partial loss of kinetoplast DNA (dyskinetoplastidy) or its complete loss (akinetoplastidy) means that the mitochondria cannot fully function, and therefore the parasite can only develop within its vertebrate host. Consequently, trypanosomes lacking a kinetoplast and those in which it is only partially functional must rely on mechanical vector transmission or sexual transmission. However, this was not an evolutionary ‘retrograde step’ because these modes of transmission broke the link with the African tsetse fly belt and allowed the parasites to spread to other countries. Dyskinetoplastidy arises naturally in wild trypanosome populations by mutation, in response to treatments with certain anti‐trypanosome drugs (e.g., triacetylbenzene hydrochloride [TBG]), and through long‐term in vitro culture.
Table 4.4 Examples of Trypanosoma species of medical and veterinary importance.
| Parasite | Transmission Group | Vector/means of transmission | Host |
|---|---|---|---|
| Trypanosoma brucei brucei | Salivaria | Tsetse flies ( Glossina spp.) | Ruminants (cause of ‘nagana’) |
| Trypanosoma brucei gambiense | Salivaria | Tsetse flies ( Glossina spp.) | Humans (cause of ‘sleeping sickness’). No important animal reservoir of infection |
| Trypanosoma brucei rhodesiense | Salivaria | Tsetse flies ( Glossina spp.) | Humans (cause of ‘sleeping sickness’). Reservoir of infection in cattle, wild game, lions, hyena etc. |
| Trypanosoma congolense | Salivaria | Tsetse flies ( Glossina spp.) | Cattle, pigs, wild game. Cause of ‘nagana’ |
| Trypanosoma evansi (Trypanosoma brucei evansi) | Salivaria | Tabanid and other biting flies (mechanical transmission) | Horse, cattle, pigs, dogs, rodents. Cause of ‘surra’ |
| Trypanosoma equinum (synonym of T. evansi ) | Salivaria | Tabanid and other biting flies (mechanical transmission) | Horses, donkeys, cattle, dogs Cause of ‘mal de caderas’ |
| Trypanosoma equiperdum (Trypanosoma brucei equiperdum) | Salivaria | Sexual transmission | Horse, asses. Cause of ‘dourine’ |
| Trypanosoma cruzi | Stercoraria | Reduviid bugs (e.g., Triatoma infestans ) | Humans (cause of Chagas Disease). Reservoir of infection in many domestic and wild animals |
| Trypanosoma theileri | Stercoraria | Tabanid flies | Cattle |
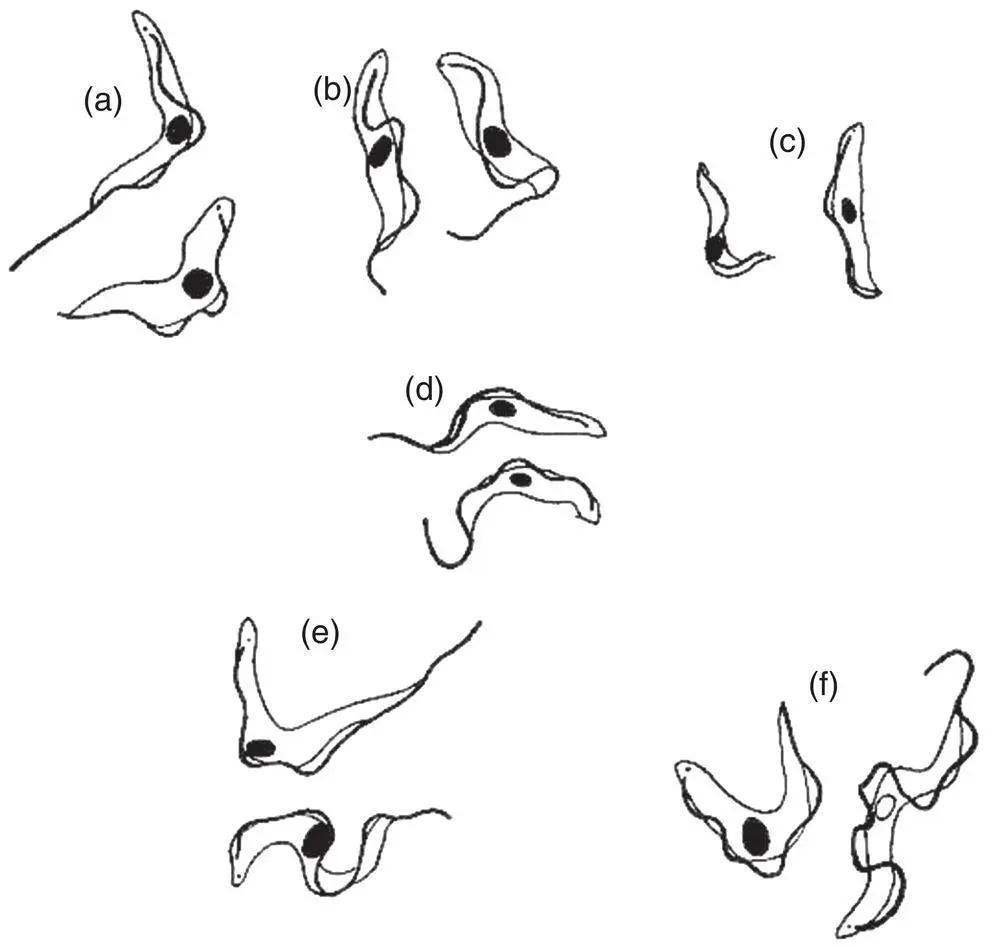
Figure 4.6 Diagrammatic representation of some Trypanosoma species. All are drawn to the same magnification. (a) Trypanosoma brucei . (b) Trypanosoma vivax . (c) Trypanosoma congolense . (d) Trypanosoma equinum . (e) Trypanosoma equiperdum . (f) Trypanosoma evansi . Note the small size and lack of a free flagellum in Trypanosoma congolense .
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Parasitology»
Представляем Вашему вниманию похожие книги на «Parasitology» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Parasitology» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.