Рафаэль Лаос-Бельтра - Том 28. Математика жизни. Численные модели в биологии и экологии.
Здесь есть возможность читать онлайн «Рафаэль Лаос-Бельтра - Том 28. Математика жизни. Численные модели в биологии и экологии.» весь текст электронной книги совершенно бесплатно (целиком полную версию без сокращений). В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Москва, Год выпуска: 2014, ISBN: 2014, Издательство: «Де Агостини», Жанр: Математика, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Том 28. Математика жизни. Численные модели в биологии и экологии.
- Автор:
- Издательство:«Де Агостини»
- Жанр:
- Год:2014
- Город:Москва
- ISBN:978-5-9774-0723-6
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Том 28. Математика жизни. Численные модели в биологии и экологии.: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Том 28. Математика жизни. Численные модели в биологии и экологии.»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Том 28. Математика жизни. Численные модели в биологии и экологии. — читать онлайн бесплатно полную книгу (весь текст) целиком
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Том 28. Математика жизни. Численные модели в биологии и экологии.», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Передача энергии между живыми организмами, населяющими экосистему, происходит в результате питания. Так как одни организмы питаются другими, образуется пищевая цепочка, которая обычно выглядит так: растение —> травоядные —» хищники —> детритофаги . В математической модели каждое звено пищевой цепи характеризуется численностью организмов, описывается циркуляция энергии и другие аспекты. Как правило, экосистемы достаточно сложны ввиду большого числа населяющих их видов и множества связей между ними, поэтому экологи стремятся упростить модели. К примеру, экосистемы грибы и бактерии могут быть объединены в рамках единой подсистемы детритофагов. Кроме того, модель экосистемы объединяется с другими моделями и становится частью итоговой общей модели. Одной из составляющих итоговой модели является окружающая среда, будь то суша, океан и т. д. Ну а для построения моделей широко используется компьютерное моделирование.
Подобным подходом к экологии мы во многом обязаны Говарду Одуму, который в 1950-е годы впервые применил для описания экосистем схемы, напоминающие схемы электрических цепей, которые сегодня лежат в основе компьютерного моделирования. Также важным вкладом Одума в науку стало объединение экологии и теории систем.
Первые математические экологические модели описывали динамику популяций. Авторы этих моделей стремились описать изменение численности популяции и ее возрастное распределение в результате взаимодействия с окружающей средой. Эти исследования берут начало в XVIII веке, когда Томас Мальтус составил модель экспоненциального роста населения, а позднее, в 1938 году, Пьер Франсуа Ферхюльст представил логистическую модель роста.
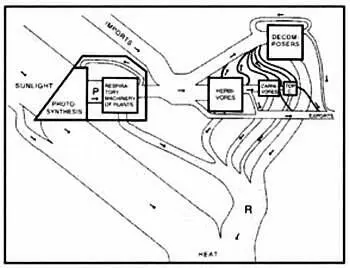
Модель, созданная на основе схем Говарда Одума. Его инновационная методология легла в основу нового способа изучения экосистем.
В 1920-е годы Вито Вольтерра и Альфред Джеймс Лотка описали две модели, которые стали настоящими столпами математической экологии. Это были модель межвидовой конкуренции и модель «хищник — жертва». Обе имели похожую структуру, однако в первой предполагалось, что рост популяций описывается логистическим уравнением, а во второй рассматривался экспоненциальный рост. Примерно двадцать лет спустя, в 1940-е годы, Патрик Лесли представил матрицу Лесли — модель структуры популяции, в которой он объединил динамику роста популяции и демографию.
В то время как в основе моделей Мальтуса, Ферхюльста и Лотки — Вольтерры лежали дифференциальные уравнения, заслуга Лесли состояла в том, что он показал применимость математических моделей, использующих матрицы.
Матричная алгебра оказалась достойным инструментом экологического и компьютерного моделирования. Матричные модели получили название BIDE (от английского Births, Immigrants, Deaths, Emigrants — «рождение, иммиграция, смерть, эмиграция»). Очевидное преимущество операций с матрицами заключалось в том, что их могли выполнять компьютеры.
В модели BIDE популяция в общем виде описывается следующим выражением:
N t+1 = N t + B + I — D — E .
* * *
ЭЛЕМЕНТАРНАЯ МОДЕЛЬ BIDE
Опишем одну из простейших экологических моделей. Допустим, что мы хотим изучить популяцию оленей, для которой известна численность самок N Jне достигших репродуктивного возраста, и число выживших самок в этой группе S Jна временном интервале от tдо t+ 1, а также численность взрослых самок N A и число выживших самок в этой группе S Aза этот же промежуток времени. Если обозначить через Р Jчисло выживших молодых самок в пересчете на каждую самку репродуктивного возраста, то численность популяции будет изменяться по следующей модели:
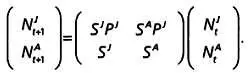
На основе этого класса элементарных моделей можно составить более сложные, к примеру модель с матрицей Лесли.
* * *
Численность популяции в момент времени t + 1 в будущем определяется ее численностью в настоящий момент времени t и совокупностью всех факторов, которые ведут к ее росту или сокращению: В — число родившихся в период времени с t по t + 1, t — число иммигрантов, присоединившихся к популяции в период времени с t по t + 1, D — число умерших в период с t по t + 1 и Е — число эмигрантов, покинувших популяцию в период с t по t + 1.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Том 28. Математика жизни. Численные модели в биологии и экологии.»
Представляем Вашему вниманию похожие книги на «Том 28. Математика жизни. Численные модели в биологии и экологии.» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Том 28. Математика жизни. Численные модели в биологии и экологии.» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.