Владимир Бердников - Эволюция и прогресс
Здесь есть возможность читать онлайн «Владимир Бердников - Эволюция и прогресс» весь текст электронной книги совершенно бесплатно (целиком полную версию без сокращений). В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Новосибирск, Год выпуска: 1991, ISBN: 1991, Издательство: Наука, Жанр: Биология, sci_popular, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Эволюция и прогресс
- Автор:
- Издательство:Наука
- Жанр:
- Год:1991
- Город:Новосибирск
- ISBN:5-02-030062-4
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Эволюция и прогресс: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Эволюция и прогресс»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Книга предназначена для биологов широкого профиля, а также всех интересующихся вопросами эволюции живых существ.
Эволюция и прогресс — читать онлайн бесплатно полную книгу (весь текст) целиком
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Эволюция и прогресс», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Ясно, что такая модель не объясняет правила Копа. Начнем ее усложнять. Быть может, мы получим искомый сдвиг распределения вправо, если зафиксируем его левую границу, т. е. примем, что вид с величиной тела меньше некоторого критического значения х* просто не может существовать. Подобное соображение выглядит вполне реалистичным. Например, для очень мелких теплокровных животных, таких как колибри у птиц и землеройки у млекопитающих, дальнейшее уменьшение размеров создало бы серьезные проблемы с поддержанием температуры тела. По-видимому, колибри, питающиеся очень калорийным нектаром, уже достигли левой границы распределения птиц. Кстати, в покое они не в состоянии поддерживать постоянную температуру.
Пусть критическое значение признака соответствует интервалу x -8 . Отсюда следует, что вид, занимающий этот самый левый интервал, может дать дочерний вид только с увеличенным размером особей, заселяя интервал х -7 . С ходом времени мы получим распределение, которое будет весьма походить на предыдущее, отличаясь от него практически только отсутствием части, расположенной слева от «поглощающего экрана» (рис. 18, б ). Естественно, с ходом времени среднее значение такого урезанного распределения будет смещаться вправо, а незначительным сдвигом его вершины (моды) можно пренебречь. Однако тщательный анализ реальных видовых распределений (подобных представленным на рис. 17) обнаруживает четкое смещение их вершины вправо, т. е. в сторону увеличения признака.
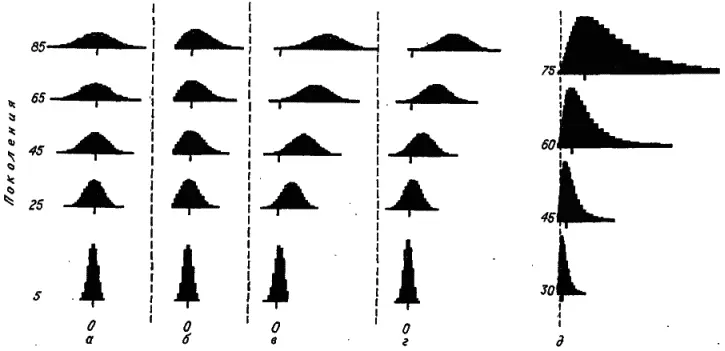
Рис. 18. Моделирование динамики видовых распределений по логарифму величины количественного признака. а — вероятность изменения признака в обоих направлениях равна и не зависит от его величины; б — те же условия, но признак не может принимать значения меньше, чем x -8; в — ортоселекция (вероятность увеличения признака в 2 раза выше вероятности его уменьшения); г — рост адаптируемости (вероятность изменения признака в обоих направлениях равна и возрастает на 0,01 с переходом в соседний интервал справа); д — те же условия, что и в случае г (признак измеряется арифметической шкалой).
Есть два пути получить динамику распределений, близкую к реальной. Первый состоит во введении асимметрии в процесс заселения смежных интервалов, второй — в ускорении заселения обоих соседних интервалов (вполне симметричного) по мере увеличения размеров особей. Первый путь — это ортоселекция, когда дочерний вид заселяет быстрее интервал справа, чем интервал слева. Если мы предположим, что вероятность заселения правого интервала в 2 раза выше, чем левого, то получим картину, представленную на рис. 18, в . Видно, что распределение быстро расползается вправо и в ту же сторону сдвигается его вершина.
Обычно современные приверженцы синтетической теории эволюции считают, что за эти макроэволюционные тенденции ответственна именно ортоселекция. Их соображения выглядят примерно так. Увеличенные размеры тела дают индивидам очевидные преимущества: их бег становится быстрее и экономичнее, они лучше защищены от хищников и, наконец, они, обладая большей мышечной силой, имеют преимущества в драках с конкурентами за спаривание. Казалось бы все логично, хотя несколько антропоморфно. Действительно, людям свойственно воспевать крупных мужчин, их, так сказать, боевые качества. Попробуем посмотреть на это с иной точки зрения.
Увеличенная масса тела затрудняет бег по пересеченной местности, к тому же крупный индивид требует для поддержания жизни больше пищи. Кроме того, с увеличением массы тела, безусловно, падает скорость реакции на внешние раздражители — весьма немаловажное обстоятельство для выживания. Едва ли существует положительная корреляция между размерами тела охотника и количеством приносимой им добычи — основы благополучия первобытного человека. Наконец, увеличенные размеры означают некоторое удлинение онтогенеза и соответственно снижение шансов достичь взрослой стадии. Таким образом, без точных оценок всех плюсов и минусов мы не в состоянии найти простую связь между размерами особи и ее приспособленностью. Единственной твердо установленной закономерностью, связывающей размеры тела животных с приспособленностью, является правило Бергмана, согласно которому среднепопуляционная масса тела у теплокровных животных одного вида увеличивается, с понижением среднегодовой температуры. Этому правилу подчиняется большинство видов птиц и млекопитающих, верно оно и для людей.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Эволюция и прогресс»
Представляем Вашему вниманию похожие книги на «Эволюция и прогресс» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.









Обсуждение, отзывы о книге «Эволюция и прогресс» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.