Владимир Бердников - Эволюция и прогресс
Здесь есть возможность читать онлайн «Владимир Бердников - Эволюция и прогресс» весь текст электронной книги совершенно бесплатно (целиком полную версию без сокращений). В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Новосибирск, Год выпуска: 1991, ISBN: 1991, Издательство: Наука, Жанр: Биология, sci_popular, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Эволюция и прогресс
- Автор:
- Издательство:Наука
- Жанр:
- Год:1991
- Город:Новосибирск
- ISBN:5-02-030062-4
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Эволюция и прогресс: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Эволюция и прогресс»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Книга предназначена для биологов широкого профиля, а также всех интересующихся вопросами эволюции живых существ.
Эволюция и прогресс — читать онлайн бесплатно полную книгу (весь текст) целиком
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Эволюция и прогресс», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Поскольку мы ввели адаптируемость как неотъемлемое свойство генома типичной особи вида, то тем самым признали, что могут быть мутации как уменьшающие, так и увеличивающие адаптируемость. Фактически мы предположили, что могут существовать мутации, изменяющие скорость появления благоприятных мутаций. На первый взгляд кажется, что для этого достаточно просто повысить мутабильность среднего гена, однако такое решение вопроса весьма сомнительно. Ведь рост мутабильности автоматически означает увеличение генетического груза, т. е. падение уровня приспособленности популяции. Наиболее перспективно выглядят мутации, повышающие долю благоприятных среди всех мутаций, возникающих в геноме за поколение. Если такие мутации, какими бы фантастичными они ни казались, действительно существуют, то они должны накапливаться в генофондах видов при браковке филетических линий по адаптируемости.
В настоящее время существуют две точки зрения на филетическое видообразование. Одна традиционная, градуалистская, впервые высказанная Дарвином, предполагает, что нет четкой границы, где один вид сменяет другой, поэтому сдвиг фенотипа средней особи идет постоянно, хотя в его скорости возможны значительные колебания. Согласно другой, высказанной С. Гоулдом и Н. Элдриджем в 1972 г., вид в течение миллионов лет не изменяется, находясь в состоянии морфологического «стазиса». Затем за относительно короткий промежуток времени происходит его превращение в новый вид — видовая трансформация. Хотя этот «краткий» миг может длиться и несколько десятков тысяч лет, все равно его длительность ничтожна в сравнении с миллионами лет стазиса.
Если верна градуалистская модель, то процесс адаптации к новым условиям длится миллионы лет, и все это время сохраняются условия для фиксации мутаций, увеличивающих адаптируемость. Пунктуалистская модель предполагает, что отбор в пользу таких мутаций действует наверняка только в период видовой трансформации, и ценные для этого момента аллели могут быть утрачены во время стазиса, если они не вносят вклад в относительную приспособленность особей. Следовательно, аллели, благоприятные для филетического выживания вида, должны быть благоприятными и в микроэволюционном смысле, т. е. повышать уровень относительной приспособленности их носителей.
Заметим, что, введя представление об адаптируемости, мы получили, как следствие, макроэволюционную тенденцию к ее росту. Эту тенденцию можно было бы интерпретировать как накопление эволюционирующей филетической группой информации о неблагоприятных факторах, действующих в ее экологическом пространстве (адаптивной зоне). Как подойти к анализу подобного рода информации, вероятно, запечатленной геномом? Скорее всего такая генетическая запись должна каким-то образом отразиться на фенотипе особей. Поэтому и тенденция к росту адаптируемости в ходе эволюции филетической группы могла бы выглядеть как долговременная тенденция к изменению в определенном направлении морфологии особей ее видов. В связи с этим встает вопрос, не являются ли хорошо известные палеонтологам долговременные тенденции отражением процесса возрастания адаптируемости.
Макроэволюционные тенденции
Крупнейший вклад палеонтологии в наши знания об эволюции живых организмов состоит в обнаружении во многих филетических группах особых очень долговременных тенденций к сдвигу в одном направлении некоторых количественных признаков. Пожалуй, самой известной из них является тенденция к увеличению размеров индивидов — так называемое правило Копа. Значительный эмпирический материал свидетельствует, что обычно зоологические таксоны крупных животных происходят от видов с относительно небольшими особями. Между прочим, род Homo происходит от гораздо более мелких обезьян.
Э. Коп объяснил свое правило как классический ламаркист (кстати, основоположник психоламаркизма). Он считал, что в основе открытой им тенденции лежит пресловутая «ростовая сила», стремящаяся под действием психики животного увеличить его размеры. Такой филогенетический рост тела протекает беспрепятственно в периоды геологического затишья и сопровождается разрастанием многих морфологических структур. Тенденции противостоит процесс вымирания гигантов и монстров при резких сменах климата.
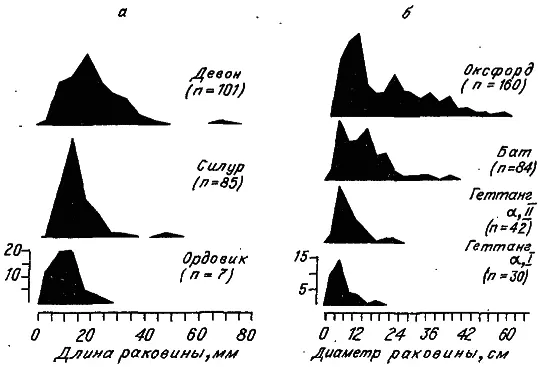
Рис. 17. Изменение видовых распределений брахиопод (а) и аммонитин (б) по размеру раковины в историческом развитии (по: [Stanley, 1973]).
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Эволюция и прогресс»
Представляем Вашему вниманию похожие книги на «Эволюция и прогресс» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.









Обсуждение, отзывы о книге «Эволюция и прогресс» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.