Brenda A. Wilson - Bacterial Pathogenesis
Здесь есть возможность читать онлайн «Brenda A. Wilson - Bacterial Pathogenesis» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Bacterial Pathogenesis
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Bacterial Pathogenesis: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Bacterial Pathogenesis»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Completely revised and updated, and for the first time in stunning full-color, Bacterial Pathogenesis: A Molecular Approach, Fourth Edition, builds on the core principles and foundations of its predecessors while expanding into new concepts, key findings, and cutting-edge research, including new developments in the areas of the microbiome and CRISPR as well as the growing challenges of antimicrobial resistance. All-new detailed illustrations help students clearly understand important concepts and mechanisms of the complex interplay between bacterial pathogens and their hosts. Study questions at the end of each chapter challenge students to delve more deeply into the topics covered, and hone their skills in reading, interpreting, and analyzing data, as well as devising their own experiments. A detailed glossary defines and expands on key terms highlighted throughout the book. Written for advanced undergraduate, graduate, and professional students in microbiology, bacteriology, and pathogenesis, this text is a must-have for anyone looking for a greater understanding of virulence mechanisms across the breadth of bacterial pathogens.
Bacterial Pathogenesis — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Bacterial Pathogenesis», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
All normal, healthy (uninfected) cells express class I major histocompatibility complex (MHC I) proteins on their surfaces, which will be discussed in more detail in the next chapter on the adaptive immune response. NK cells can distinguish between uninfected and infected cells based on the presence or absence of MHC I, respectively. The complex of MHC I bound with a self antigen (small peptides that are derived from the host cell and not from a foreign invading pathogen) is recognized by an MHC I-specific inhibitory receptor on the NK cell surface, which then sends a signal inside the NK cell that halts activation of the cytotoxic response (release of granules) stimulated by the activating ligand-receptor interactions.
By contrast, infected cells often express much less MHC I on the surface than normal cells do, and those MHC I molecules that are on the surface are more likely to have foreign antigens (rather than self antigens) bound. Complexes of MHC I bound to foreign antigens are recognized by CTLs of the adaptive immune system (see chapter 4). The absence of self antigen-bound MHC I on the surface means that the NK cell activation response is not blocked by its inhibitory receptors, and this allows the NK cell to proceed with its attack on the infected cell.
The Lymphatic System
Lymph, the fluid that moves through the lymphatic system, is also monitored and protected by phagocytes. Lymphatic vessels are tubes comprised of overlapping endothelial cells. These tubes are organized into a network similar in complexity to the circulatory system that carries blood to all parts of the body ( Figure 3-4). The role of the lymphatic system is to prevent excess buildup of fluid in tissues and to recycle blood proteins. Under healthy conditions, blood fluids and proteins leak from blood vessels in the capillary beds to feed cells, donate oxygen, and remove carbon dioxide. These fluids also provide protective blood proteins such as complement components (see below), cytokines, and chemokines. Blood vessels, however, do not readily reabsorb the fluids that leak from them. Rather, these fluids are taken care of by the lymphatic vessels. Lymphatic endothelial cells are tethered to the muscle bed, so when the level of fluids in an area becomes elevated and pressure causes muscle cells to separate, the overlapping endothelial cells of the lymphatic vessels are pulled apart. This opening allows fluid from the surrounding area to enter the lymphatic vessels, where it is then returned to central holding areas such as the thoracic duct and eventually returned to the bloodstream.
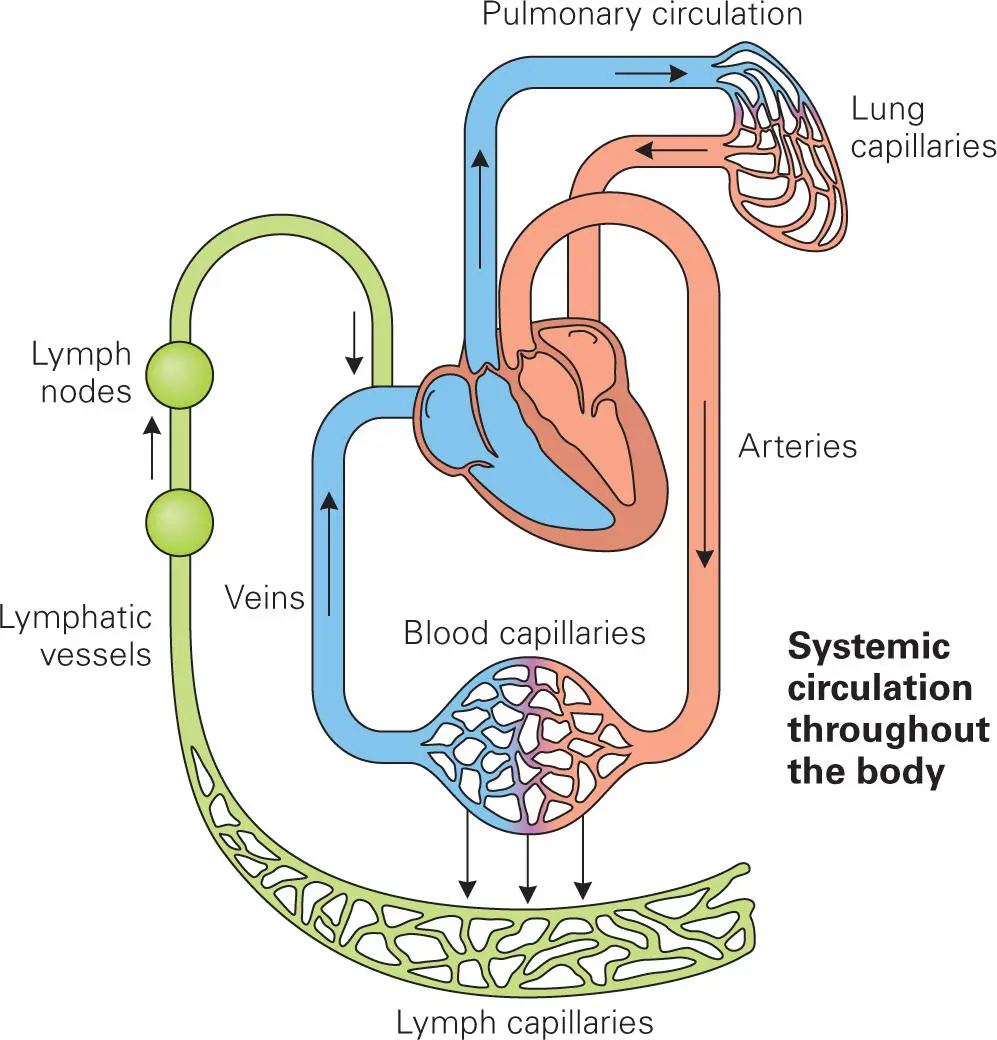
Figure 3-4. The lymphatic system.
The inflammatory response to bacterial infection creates a buildup of fluid in tissue, leading to opening of the lymphatic vessels. We experience the accumulation of blood fluids as swelling and tightness that appears around infected wounds or sites of infection. In these situations, bacteria can enter the lymphatic vessels. In order to prevent these bacteria from spreading throughout the body, the lymph must first be cleansed of these bacteria before it reenters the bloodstream. This task is accomplished by lymph nodes located at strategic points along the lymphatic vessels.
Lymph nodes are major sites distributed throughout the body that are linked to lymphatic vessels and contain macrophages, DCs, T cells, and B cells. Analogous to how the liver and kidneys filter toxic substances from circulation, the lymph nodes function to filter foreign entities, such as pathogens. The spleen and tonsils are other large lymphoid tissues that function similarly to lymph nodes but filter blood instead of lymph. In chapter 4, we will also learn that macrophages and DCs in the lymph nodes act as antigen-presenting cells and stimulatory cells that potentiate the adaptive defense response against bacteria. Thus, the lymph nodes not only serve to sterilize lymph but also act as sites where the adaptive defense system is alerted.
Bacteria that enter a lymph node are usually killed by the macrophages and DCs, but there are bacteria that are able to evade this fate. Indeed, some of the most dangerous pathogens are those that can survive and multiply in the lymph nodes. An example of such a pathogen is Yersinia pestis, the cause of bubonic plague (also known as the Black Death). Y. pestis growing in lymph nodes creates an inflammatory response so intense that it causes the lymph nodes to become grossly distended (swollen), producing the so-called buboes that give bubonic plague its name. A less pronounced, but still detectable, enlargement or swelling of lymph nodes (lymphadenopathy) likewise occurs during many types of bacterial infections and serves as a diagnostic sign of infection.
How Phagocytes Recognize and Respond to Bacteria
For many years, the interaction between PMNs and invading bacteria remained mysterious. With the discovery of PAMPs and their host cell receptors, this interaction has been a focus of intense research in recent years. Much of this research has focused on components of Gram-negative bacteria, such as the cell surface component lipopolysaccharide (LPS) (see Box 1-4) and flagellin, a protein involved in bacterial cell motility. Gram-positive bacteria also trigger the same type of PMN activation through lipoteichoic acid (LTA), the anionic polymer found in the cell walls of these bacteria (see Box 1-4). Peptidoglycan (PG) fragments from both Gram-positive and Gram-negative bacteria, as well as certain types of DNA (e.g., CpG-rich DNA) and RNA, can also act as triggers.
PAMPs were discovered in an unusual way. Scientists studying the fruit fly, Drosophila melanogaster, identified a transmembrane receptor protein called Toll that was required for resistance to fungal infections. Oddly enough, only one of the Toll receptors in Drosophila is linked to a defense against infection; the others are involved in various aspects of fly development. Shortly thereafter the mammalian equivalent of Toll was discovered—hence the name “Toll-like receptor (TLR),”—and shown to be important for immunity. To date, thirteen mammalian TLR paralogs (genes found in different species that are traceable to a shared ancestry) have been described and have been shown to modulate cellular responses to pathogens. Most mammals have ten to thirteen different TLRs (humans have ten functional TLRs).
Although there are many different members of the TLR family, one common set of them is evolutionarily conserved in a wide range of organisms. This common set of diverse TLRs may reflect the fact that although all Gram-negative and Gram-positive bacteria have surface “signatures” in common, there are also numerous differences. The different TLRs in various combinations (through homo- and heterodimerization) and coupling with different downstream adaptor molecule combinations provide the flexibility to respond to differences between invading bacteria without responding specifically to each particular bacterium, as the antibody-based adaptive immune system does. For example, TLR4 is a receptor for LPS from most Gram-negative bacteria, but sometimes a complex of TLR1/TLR2 or TLR2/TLR6 is the receptor for certain pathogens that have modified LPS structures or surface lipoproteins. TLR9 responds to DNA containing stretches of unmethylated CpG sequences, which is not usually found in mammalian DNA. Double-stranded RNA activates TLR3 and TLR8, while single-stranded RNA activates TLR7, and bacterial flagellin activates TLR5. The ligand for TLR10 is not yet known. Mice have a TLR11 that recognizes parasites and uropathogenic Escherichia coli, but in humans the gene encoding TLR11 has multiple stop codons that prevent expression of a functional protein.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Bacterial Pathogenesis»
Представляем Вашему вниманию похожие книги на «Bacterial Pathogenesis» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Bacterial Pathogenesis» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.