Keith N. Frayn - Human Metabolism
Здесь есть возможность читать онлайн «Keith N. Frayn - Human Metabolism» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Human Metabolism
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Human Metabolism: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Human Metabolism»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
(formerly
) offers a current and integrated review of metabolism and metabolic regulation. The authors explain difficult concepts in clear and concise terms in order to provide an accessible and essential guide to the topic. This comprehensive text covers a wide range of topics such as energy balance, body weight regulation, exercise, and how the body copes with extreme situations, and illustrates how metabolic regulation allows the human body to adapt to many different conditions.
This fourth edition has been revised with a new full colour text design and helpful illustrations that illuminate the regulatory mechanisms by which all cells control the metabolic processes necessary for life. The text includes chapter summaries and additional explanatory text that help to clarify the information presented. In addition, the newly revised edition includes more content on metabolic pathways and metabolic diseases. This important resource:
Is a valuable tool for scientists, practitioners and students across a broad range of health sciences including medicine, biochemistry, nutrition, dietetics, sports science and nursing Includes a full colour text filled with illustrations and additional diagrams to aid understanding Offers a companion website with additional learning and teaching resources. Written for students of medicine, biochemistry, nutrition, dietetics, sports science and nursing,
has been revised and updated to provide a comprehensive review of metabolism and metabolic regulation.
Human Metabolism — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Human Metabolism», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
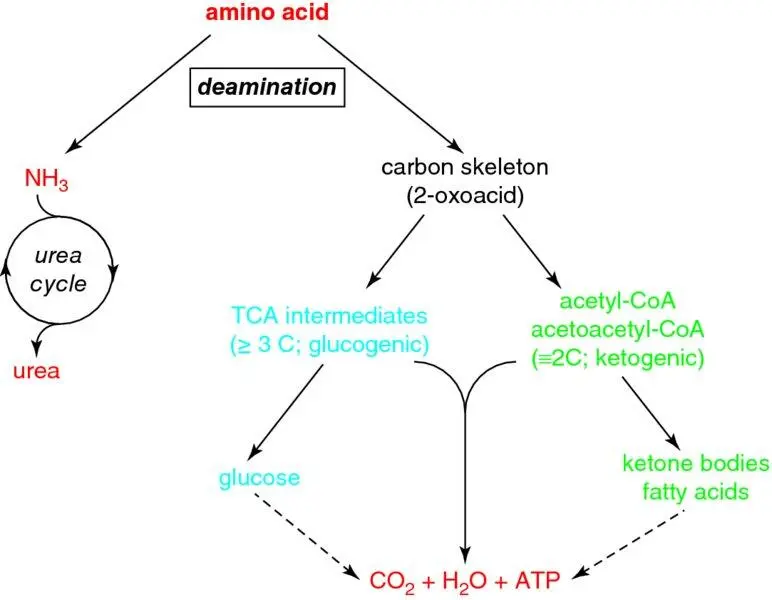
Figure 1.19 Metabolism of amino acids. To be metabolised, amino acids must first be deaminated to remove the nitrogen (amino group) from the central α carbon. This leaves the carbon skeleton (the corresponding 2-oxoacid [α-ketoacid]) which undergoes metabolism in the common metabolic pool, by a route which depends on its structure, ultimately producing energy. The amino group becomes ammonia, some of which can be excreted directly in the urine, but most of which must be detoxified by the urea cycle.
1.3.4.2 Amino acid-nitrogen disposal
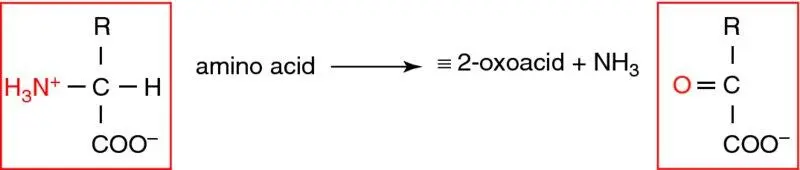
Deamination of amino acids is achieved by two types of reaction which function in a complementary manner. The first is transamination , in which the amino group from one amino acid is transferred to another 2-oxoacid (carbon skeleton), forming its corresponding amino acid (i.e. amino acid-1 + 2-oxoacid-2 ↔ 2-oxoacid-1 + amino acid-2). The enzymes responsible for transamination reactions are aminotransferases ( transaminases ), all of which contain pyridoxal phosphate, a derivative of vitamin B6, in their active centres ( Figure 1.20). Aminotransferases are widespread in most tissues and are near-equilibrium, and hence readily reversible. Each is specific for a limited number of amino acids, but most utilise 2-oxoglutarate (also called α-ketoglutarate) as the amino (N) acceptor (the ‘2-oxoacid-2’ in Figure 1.20), producing the carbon backbone of the donor amino acid together with glutamate (amino acid + 2-oxoglutarate → 2-oxoacid + glutamate). Hence 2-oxoglutarate and glutamate are central to amino acid catabolism as these reactions ‘funnel’ the various amino acids into glutamate (and note that 2-oxoglutarate is common to the TCA cycle, and its utilisation by this pathway depletes TCA cycle intermediates – cataplerosis). Alanine aminotransferase (ALT) transfers the amino group of alanine to 2-oxoglutarate, forming pyruvate and glutamate. The pyruvate can be used for gluconeogenesis (see below), e.g. during starvation. Alanine is a key transport form of amino acid carbon and nitrogen, hence this enzyme is important for inter-tissue amino acid flux. Aspartate aminotransferase (AST) transfers the amino group of aspartate to 2-oxoglutarate, forming oxaloacetate and glutamate – however, this enzyme usually works in the reverse direction, its function being to convert glutamate (derived from amino acid funnelling, above) into aspartate, which is required to donate a second urea N-atom to the urea cycle. Since ALT and AST are both intracellular enzymes and widespread, necrosis of many tissues, including liver, causes them to increase in plasma; they are commonly used to diagnose hepatocellular damage (so-called ‘ liver function tests ’).
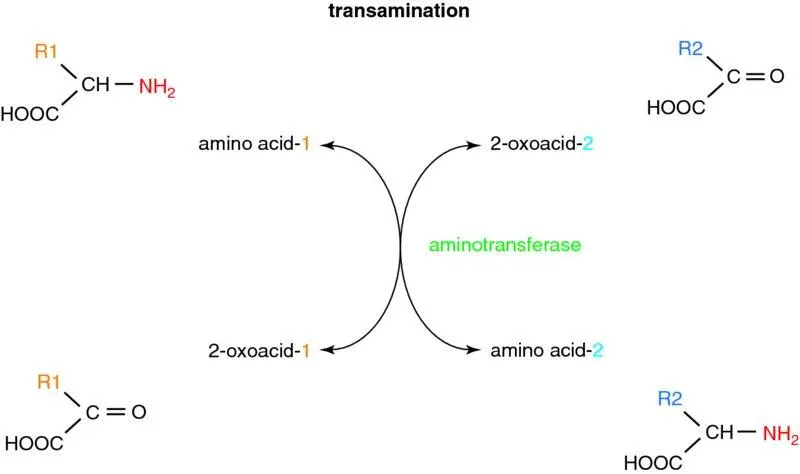
Figure 1.20 Transamination reactions. Transamination involves the transfer of an amino group from the α-carbon of an amino acid to a recipient 2-oxoacid (α-ketoacid), forming its corresponding 2-oxoacid and generating an amino acid. These reactions are catalysed by aminotransferase enzymes, all of which are readily reversible. Although no net deamination occurs, these reactions allow the amino groups of all amino acids to be ‘funnelled’ into key amino acids prior to net deamination and hence metabolism.
The second type of reaction responsible for amino acid deamination is oxidative deamination . Since most amino acids have been funnelled (deaminated) into glutamate by transamination, glutamate is the only amino acid that undergoes direct, oxidative, deamination, by glutamate dehydrogenase, regenerating 2-oxoglutarate and producing ammonia (NH 3). Unusually for a highly regulated enzyme, glutamate dehydrogenase is reversible, and can use NAD +/NADH or NADP +/NADPH as electron carriers. In the ‘forward’ direction of deamination (catabolic, amino acid breakdown), it uses NAD +, but in the ‘reverse’ direction of amination of 2-oxoglutarate to glutamate (anabolic, amino acid synthesis) it uses NADPH, reflecting the different roles of these cofactors as redox energy carriers in different metabolic states ( Box 1.8; for simplicity, ionisation states are not always shown as they would be at physiological pH; ammonium ion shown here [NH 4 +] may be considered the same as ammonia). Hence, aminotransferases (transamination) and glutamate dehydrogenase (oxidative deamination) work together to produce ammonia for detoxification to urea in the urea cycle, and carbon skeletons for further intermediary metabolism ( Figure 1.21).
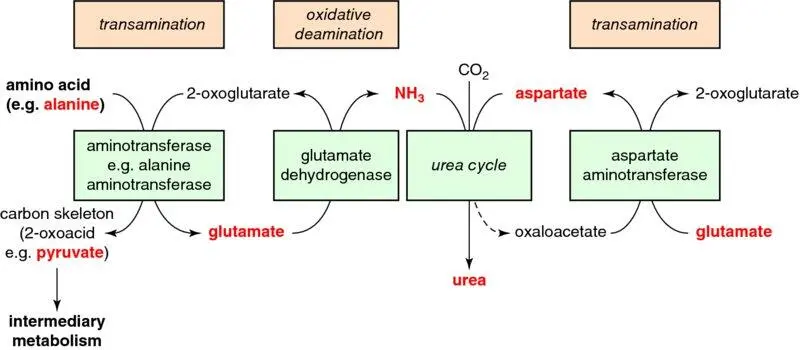
Figure 1.21 Deamination of amino acids. By linking transamination reactions to oxidative deamination, and then to the urea cycle, all amino acids can be efficiently deaminated, their carbon backbones (2-oxoacids) going on to further metabolism for energy production, and the amino group being safely detoxified by the urea cycle.
Box 1.8 Deamination
transamination is a type of deamination but does not remove net N
presence of α-amino group prevents oxidative breakdown
therefore α-amino group must be removed before catabolism can proceed
the nitrogen can be incorporated into other compounds or excreted in the urine
different types of deamination but oxidative deamination is quantitatively the most important
| • oxidative | glutamate dehydrogenase |
| • non-oxidative | serine & threonine: hydroxyl in side chain |
| • hydrolytic | asparagine & glutamine: N in side chain |
glutamate is the only amino acid that undergoes oxidative deamination (glutamate dehydrogenase)
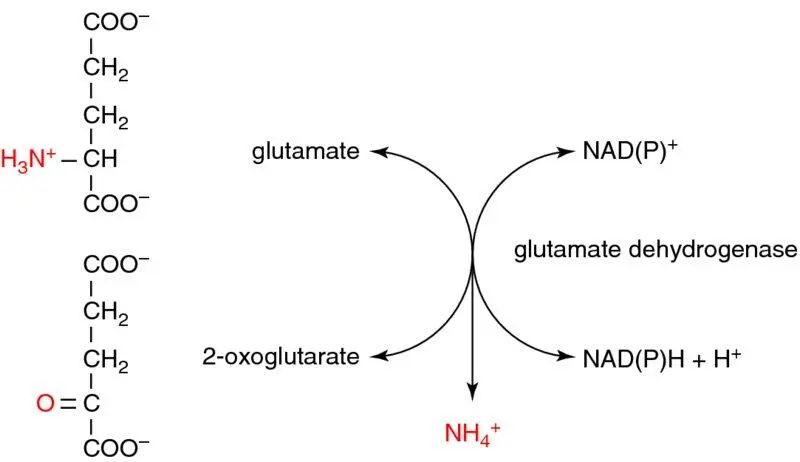
mostly occurs in liver and kidney
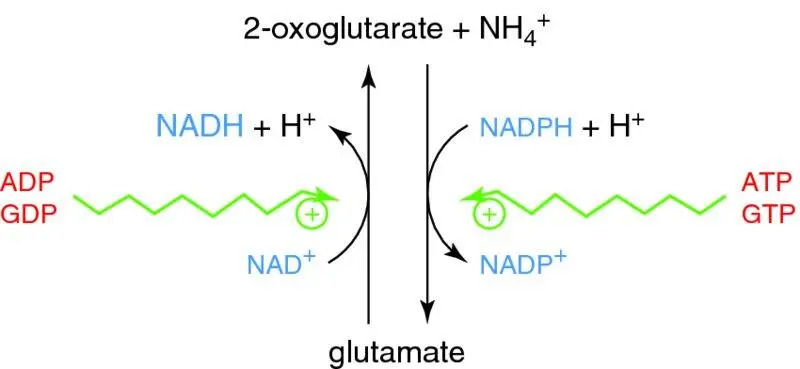
unusually can use either NAD+ or NADP+ as coenzymeNAD+ used mostly in oxidative deaminationNADP+ used mostly in reductive amination
direction of reaction depends on substrate availability (& hence metabolic state)
allosteric regulation (unusually for a readily reversible reaction):
The urea (ornithine) cycle occurs in the liver. Urea (CO·(NH 2) 2) contains two nitrogen atoms: one derives from ammonia (oxidative deamination of glutamate), the other from aspartate (transamination, also of glutamate, by AST) ( Figure 1.21): the body excretes nitrogen with minimal carbon (and energy) loss. Because urea is very water-soluble, much nitrogen waste can be excreted for relatively little water loss, an important adaptation in terrestrial animals. Urea lacks toxicity at physiological concentrations; it is (neuro)toxic only in extremely high concentrations, for example those seen in untreated renal failure, but considerably less so than ammonia.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Human Metabolism»
Представляем Вашему вниманию похожие книги на «Human Metabolism» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Human Metabolism» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.