Tina M. Henkin - Snyder and Champness Molecular Genetics of Bacteria
Здесь есть возможность читать онлайн «Tina M. Henkin - Snyder and Champness Molecular Genetics of Bacteria» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Snyder and Champness Molecular Genetics of Bacteria
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Snyder and Champness Molecular Genetics of Bacteria: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Snyder and Champness Molecular Genetics of Bacteria»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Although the text is centered on the most-studied bacteria,
and
, many examples are drawn from other bacteria of experimental, medical, ecological, and biotechnological importance. The book's many useful features include
Text boxes to help students make connections to relevant topics related to other organisms, including humans A summary of main points at the end of each chapter Questions for discussion and independent thought A list of suggested readings for background and further investigation in each chapter Fully illustrated with detailed diagrams and photos in full color A glossary of terms highlighted in the text While intended as an undergraduate or beginning graduate textbook, Molecular Genetics of Bacteria is an invaluable reference for anyone working in the fields of microbiology, genetics, biochemistry, bioengineering, medicine, molecular biology, and biotechnology.
"This is a marvelous textbook that is completely up-to-date and comprehensive, but not overwhelming. The clear prose and excellent figures make it ideal for use in teaching bacterial molecular genetics."—
, University of Washington
Snyder and Champness Molecular Genetics of Bacteria — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Snyder and Champness Molecular Genetics of Bacteria», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
These systems are required for the secretion of proteins involved in pathogenesis and symbiosis, but they are also found in marine and soil bacteria, where they may have roles in biofilm formation and cell-to-cell communication. There can be little doubt of the importance of these systems, since mutations in their genes can have dramatic phenotypes, but many questions remain about their structure and function.
Protein Secretion in Bacteria That Lack an Outer Membrane
So far, we have limited our discussion of protein transport to the mechanisms used by bacteria that contain an outer lipid bilayer membrane (commonly referred to as Gram-negative bacteria). However, a large group of bacteria, notably the Firmicutes and Actinobacteria , lack an outer membrane (and are often referred to as the Grampositive bacteria). Organisms in this group have evolved different types of secretion systems.
INJECTOSOMES
Pathogenic bacteria that lack the secretion systems described above require alternative mechanisms to translocate virulence effectors into eukaryotic cells. Some AB-type toxins, such as diphtheria toxin, and clostridial neurotoxins, such as botulinum toxin, are self-translocating, similar to the autotransporter systems described above. However, some bacteria, such as Streptococcus pyogenes , inject a virulence effector into the mammalian target cell by a mechanism functionally analogous to the T3SS (see Madden et al., Suggested Reading). This has been named an injectosome to distinguish it from the standard type III injectisome (note the different spellings). As shown in Figure 2.41, the function of the injectosome requires cotranslational translocation of the effector by the Sec-dependent secretion system across the bacterial membrane before it translocates the effector through the cell wall and directly across the membrane of the eukaryotic target cell.
Sortases
In bacteria that lack a bilipid outer membrane, the cell wall is available to the external surface of the cell. These bacteria take advantage of this by attaching some proteins directly to their cell wall so that the proteins will be exposed on the cell surface. Proteins destined for covalent attachment to the outer surface of the cell wall are the targets of a type of cell wall-sorting enzyme called a sortase. The general sortase, sortase A (StrA), sometimes called the housekeeping sortase, may attach as many as 40 different proteins to the cell wall. Others are more specific for certain proteins.
A sortase is able to create covalent attachments between peptides by catalyzing a transpeptidation reaction. Figure 2.42illustrates a typical StrA pathway. Surface proteins that are StrA targets contain an N-terminal signal sequence and a 30- to 40-residue C-terminal sorting signal, which is composed of a pentapeptide cleavage site, LPXTG, and a hydrophobic domain that together constitute the cell wall-sorting signal (Cws) ( Figure 2.42A). The N-terminal signal sequence of the sortase target protein directs the protein to the membrane translocase, where the signal sequence is removed ( Figure 2.42B). After the protein has been translocated across the cytoplasmic membrane, the sortase cuts between the threonine (T) and glycine (G) in the pentapeptide sorting signal and then covalently links the carboxyl group of the threonine to a specific cysteine in the sortase C terminus. The sortase then attaches it to the terminal amino acid in the lipid II interlinking peptide that links two peptide crosslinks in the cell wall. When the N -acetylmuramic acid (MurNac) in this lipid II is incorporated into the cell wall, the protein becomes covalently attached to the cell wall.
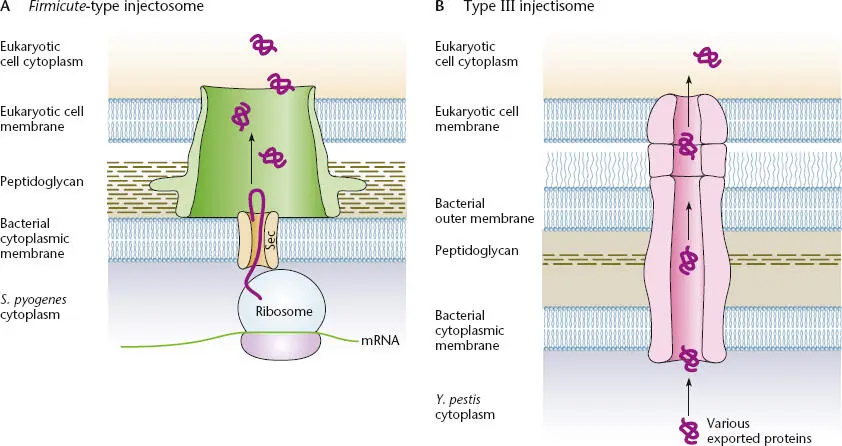
Figure 2.41 Comparison of the Firmicute-type injectosome (A)and the type III injectisome (B). Modified from Tweten RK, Caparon M, in Waksman G, Caparon M, Hultgren S (ed), Structural Biology of Bacterial Pathogenesis (ASM Press, Washington, DC, 2005).
Sortase subfamilies are defined based on their taxonomic distributions and differences in the sorting signal and the peptide to which it can be attached (see Hendrickx et al., Suggested Reading). A particularly interesting example is the assembly of pili from their subunits on the cell surfaces. Pili are composed of one major subunit, often called pilin, and one or more minor subunits. In the simplest example, pili composed of pilin and only one other subunit, the minor subunit at the tip is attached to the first pilin subunit by a specialized sortase, and this first subunit is then attached to a number of other pilin subunits by the same specialized sortase. Finally, the last subunit is attached to the cell wall by the housekeeping sortase A, which attaches most other proteins to the cell wall. This makes for a very stable pilus and secures its attachment to the cell wall.
Regulation of Gene Expression
The previous sections have reviewed how a gene is expressed in the cell, from the time mRNA is transcribed from the gene until the protein product of the gene reaches its final destination in or outside the cell and has its effect. In general, genes are expressed in the cell only when the product is needed by the cell and then only as much as is required to make the amount of product needed by the cell. This saves energy and prevents the products of different genes from interfering with each other. The process by which the output of genes is changed depending on the state of the cell is called the regulation of gene expressionand can occur at any stage in the expression of the gene (see chapter 11).
Genes whose products regulate the expression of other genes are called regulatory genes. The product of a regulatory gene can either inhibit or stimulate the expression of a gene. If it inhibits expression, the regulation is negative; if it stimulates expression, the regulation is positive. Some regulatory gene products are both positive and negative regulators depending on the situation. The product of a regulatory gene can regulate the expression of only one other gene, or it can regulate the expression of many genes. The set of genes regulated by the same regulatory gene product is called a regulon. If a gene product regulates its own expression, it is said to be autoregulated. We discuss the molecular mechanisms of regulation of gene expression in much more detail in chapter 11, but in this chapter, we briefly review some basic concepts needed to understand the following chapters.
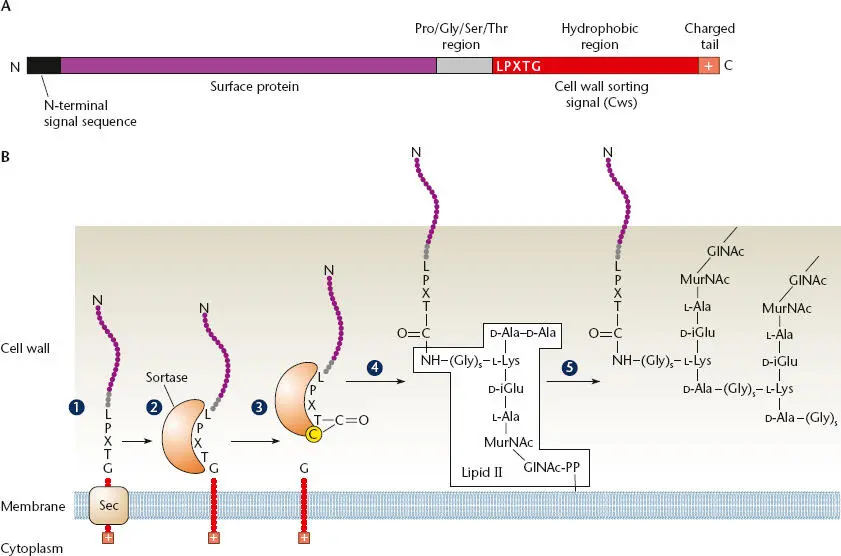
Figure 2.42 The sortase A pathway. (A)Typical sortase substrate. The protein is composed of an N-terminal signal peptide and a C-terminal cell wall-sorting signal (Cws). The Cws contains a conserved LPXTG motif followed by a hydrophobic stretch of amino acids and positively charged residues at the C terminus. (B)Model for the cell wall sortase A pathway in Staphylococcus aureus . (1)The full-length surface protein precursor is secreted through the cytoplasmic membrane via an N-terminal signal sequence. (2)A charged tail (+) at the C terminus of the protein may serve as a stop transfer signal. Following cleavage of this secretion signal, a sortase enzyme cleaves the protein between the threonine and glycine residues of the LPXTG motif, forming a thioacyl-enzyme intermediate to a specific cysteine in the sortase (3). It is then attached to the free amine of the five-glycine cross-bridge of lipid II (4)before transfer into the cell wall (5). The Pro-Gly-Ser-Thr region may help it through the thick cell wall so that it is expressed on the cell surface. PP is the site of MurNAc pentapeptide attachment to bactoprenyl in the membrane in lipid II. Modified from Connolly KM, Clubb RT, in Waksman G, Caparon M, Hultgren S (ed), Structural Biology of Bacterial Pathogenesis (ASM Press, Washington, DC, 2005).
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Snyder and Champness Molecular Genetics of Bacteria»
Представляем Вашему вниманию похожие книги на «Snyder and Champness Molecular Genetics of Bacteria» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.











Обсуждение, отзывы о книге «Snyder and Champness Molecular Genetics of Bacteria» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.