Jane Flint - Principles of Virology
Здесь есть возможность читать онлайн «Jane Flint - Principles of Virology» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.
- Название:Principles of Virology
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Principles of Virology: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Principles of Virology»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Volume I: Molecular Biology
Volume II: Pathogenesis and Control
Principles of Virology, Fifth Edition
Principles of Virology — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Principles of Virology», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
The first steps in virus attachment are governed largely by the probability of a random collision between a virus particle and a cell, and therefore by the concentrations of free particles and host cells. The rate of attachment can be described by the equation
dA / dt = k [ V ][ H ]
where A is attachment, t is time, [ V ] and [ H ] are the concentrations of virus particles and host cells, respectively, and k is a constant that defines the rate of the reaction. It can be seen from this equation that if a mixture of viruses and cells is diluted after a period sufficient for adsorption, subsequent binding of particles is reduced greatly. For example, a 100-fold dilution of the virus and cell mixture reduces the attachment rate 10,000-fold (i.e., 1/100 × 1/100).
The initial association of virus particles with cells is probably via electrostatic forces, as it is sensitive to low pH or high concentrations of salt, but higher-affinity binding relies mainly on hydrophobic and other short-range forces. These interactions are usually aided by the extracellular matrix. Extracellular matrices hold the cells and tissues of the body together and are made up of two main classes of macromolecules: glycosaminoglycans (such as heparan sulfate and chondroitin sulfate), which are unbranched polysaccharides made of repeating disaccharides; and fibrous proteins with structural (collagen and elastin) or adhesive (fibronectin and laminin) functions ( Fig. 5.1).
PRINCIPLES Attachment and entry
Viruses that infect animals produce particles that are too large to diffuse across the plasma membrane, and thus entry must be an active process.
Virus particles bind to receptors on their host cells to initiate entry.
The cell receptor is a major determinant of the host range and tissue tropism of the virus.
Virus particles may bind multiple distinct receptors, and individual cellular proteins may be receptors for multiple viruses.
Enveloped virus particles bind receptors via their transmembrane glycoproteins; nonenveloped virus particles bind via the capsid surface or projections from the capsid.
Many virus particles enter host cells by the same cellular pathways used to take up macromolecules.
The mechanism of entry of a particular virus may depend on the nature of the target cell.
Binding of virus particles to cell receptors may activate signaling pathways that facilitate virus entry and movement, or produce cellular responses that enhance virus propagation and/or govern pathogenesis.
Some virus particles uncoat at the plasma membrane, while others do so at intracellular vesicles.
The cytoskeleton and associated motors facilitate movement of viral and subviral particles within the infected cell.
Nuclear import of viral components can occur either via the nuclear pore complex or during cell division, when the nuclear membrane breaks down.
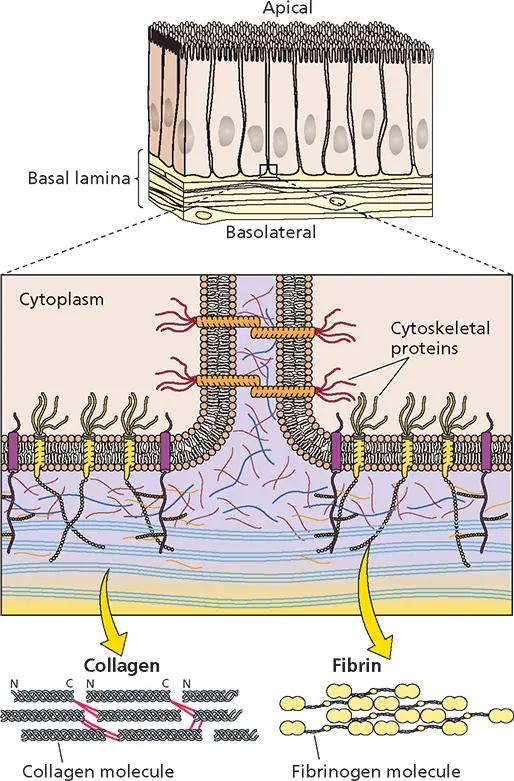
Figure 5.1 Architecture of cell surfaces. Example of epithelial cells with apical and basolateral surfaces noted. Cell surface molecules contribute to cell-cell adhesion and attachment to the extracellular matrix mainly composed of glycosaminoglycans (purple), fibrin (yellow), and collagen (blue). Tight junctions are formed by transmembrane proteins (orange) to regulate passage of molecules between cells.
The initial nonspecific interactions have to result in binding to one or more virus receptor(s) for entry to proceed. Although the affinityof a receptor for a single virus particle is typically low, the presence of multiple receptor-binding sites on the virus particle and the fluid nature of the plasma membrane allow engagement of multiple receptors. Consequently, the avidity(the strength conferred by multiple interactions) of virus particle binding to cells is typically very high. Binding can usually occur at 4°C (even though entry does not) as well as at body temperature (e.g., 37°C). Infection of cultured cells can therefore be synchronized by allowing binding to take place at a low temperature and then shifting the cells to a physiological temperature to allow the initiation of subsequent steps.
Binding of viral proteins to a specific receptor induces conformational changes in the virus particle that lead to membrane fusion or penetration. Binding might also trigger the transmission of signals by the receptor that cause virus uptake or initiation of receptor-mediated endocytosis to bring the bound particles into these pathways. Receptors for viruses comprise a variety of cell surface proteins, carbohydrates, and lipids, all with functions in the cell unrelated to virus entry. Virus receptors that have been identified include immunoglobulin-like proteins, ligand-binding receptors, glycoproteins, ion channels, gangliosides, carbohydrates, proteoglycans, and integrins. The receptor may be the only cell surface molecule required for entry into cells, or additional cell surface molecule(s), or coreceptor(s), may be needed ( Box. 5.1). Different receptors may serve for virus entry in diverse cell types (e.g., herpesviruses), and unrelated viruses may bind to the same receptor (e.g., the Coxsackievirus and adenovirus receptor).
The receptor is the main determinant of the host rangeof a virus, i.e., its ability to infect a particular animal or its cells. For example, poliovirus infects primates and primate cells in culture but not mice or mouse cells in cultures. Mouse cells synthesize a protein that is homologous to the poliovirus receptor in primates, but sufficiently different that poliovirus cannot attach to it. In this example, the poliovirus receptor is thedeterminant of poliovirus host range. However, production of the receptor in a particular cell type does notensure that virus reproduction will occur. Some primate cell cultures produce the poliovirus receptor but cannot support virus re-production; the cells are susceptible but not permissive. The restriction of viral reproduction in these cells is most probably due to a block in viral reproduction beyond the attachment step. Receptors can also be determinants of tissue tropism, the predilection of a virus to invade and reproduce in a particular cell type. However, there are many other determinants of tissue tropism. For example, the sialic acid residues on membrane glycoproteins or glycolipids, which are receptors for influenza virus, are found on many tissues, yet viral reproduction in the host is restricted as essential components for reproduction are missing from these cells. Additionally, passage of viruses in the laboratory can alter their receptor specificity. When receptors are rare, viruses that can bind to the more abundant glycosaminoglycans are readily selected. For example, adaptation of Sindbis virus to cells in culture has led to the selection of variants that bind heparan sulfate. Nevertheless, the majority of viral receptors and coreceptors identified experimentally have proven to fulfill these functions in vivo .
Many receptor molecules can move in the plasma membrane, leading to the formation of microdomains that differ in composition. Bound virus particles may therefore localize to specialized areas of the plasma membrane such as lipid rafts, subdomains rich in cholesterol and glycosphingolipids; caveolae, small invaginations formed by caveolin oligomers; or clathrin-coated pits. Such areas may promote endocytosis of receptor-bound virus particles by distinct pathways.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Principles of Virology»
Представляем Вашему вниманию похожие книги на «Principles of Virology» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Principles of Virology» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.